|
Vertebrates, Kenneth V. Kardong, 1998, ISBN 0-697-28654-1, McGraw-Hill, p 194
In use April 2000 2nd - 4th year biology at McMaster University. |
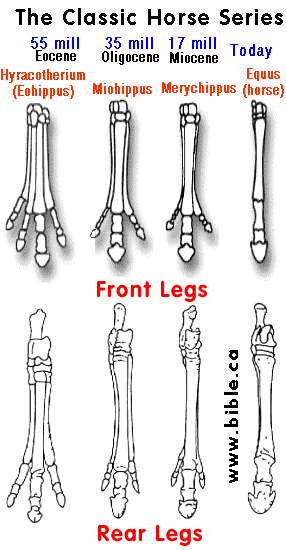
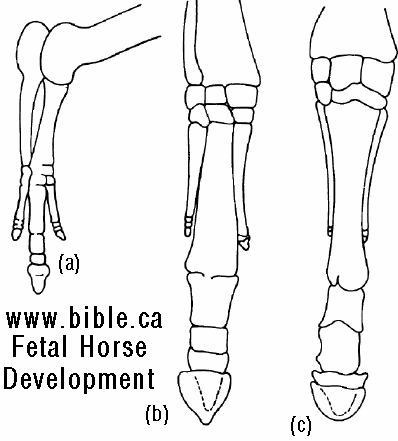
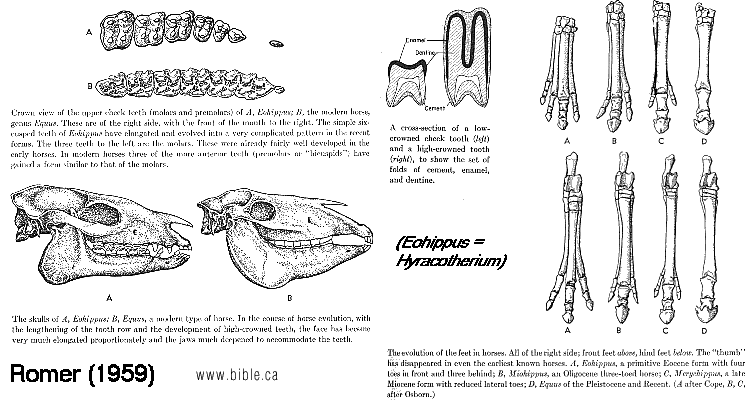
It is tempting to interpret other specialized structures in a similar light. Among modern horses, only a single toe (the middle or third toe) persists on each leg to form the functional digit (figure 5.42a). However, ancestral horses, such as Protorohippus, had four toes on the front foot and three on the back. Occasionally, modern horses develop vestiges of these old second and fourth toes (figure 5.42b-d). When this occurs, we get a glimpse Of the underlying developmental pat-tern that produces the foot. The reduction of toes in horses enjoyed adaptive favor because it contributed to locomotor performance. Literally hundreds of structural changes occurred in bones, muscles, ligaments, nerves, and blood vessels from the four (or five) toed ancestors to the single-toed modern horses. If this, like the evolution of limblessness in some lizards, was based on a narrowing of the underlying developmental pattern, then these hundreds of changes could be accomplished with relatively few gene mutations. (Vertebrates, Kenneth V. Kardong, 1998, ISBN 0-697-28654-1, McGraw-Hill, p 194)
FIGURE 5.42 (above) Extra toes in modern horses. (a) Modern horses have only one enlarged digit on each foot, a single toe. The one toe evolved from ancestors with three or four toes. During the course of their evolution, the peripheral toes IV, II, and I were lost and the central toe (III) emphasized. (b, c) On rare occasions, however, these "lost" toes or their remnants reappear, testifying to the lingering presence of the underlying ancestral developmental pattern. (d) On rare occasions, modern horses, such as the one illustrated, exhibit additional toes. Such toe remnants in modern horses apparently represent the partial reemergence of an ancient ancestral pattern. For more information on extra toes in modern horses, see Gould, S. J. 1983. Hen's teeth and horse's toes. Further reflections in natural history. New York-.W. W. Norton. (Vertebrates, Kenneth V. Kardong, 1998, ISBN 0-697-28654-1, McGraw-Hill, p 195) |
|
Evolutionary Biology, Douglas J. Futuyma, Third Edition, 1998 ISBN: 0-87893-189-9
In use April 2000 2nd - 4th year biology at McMaster University. |
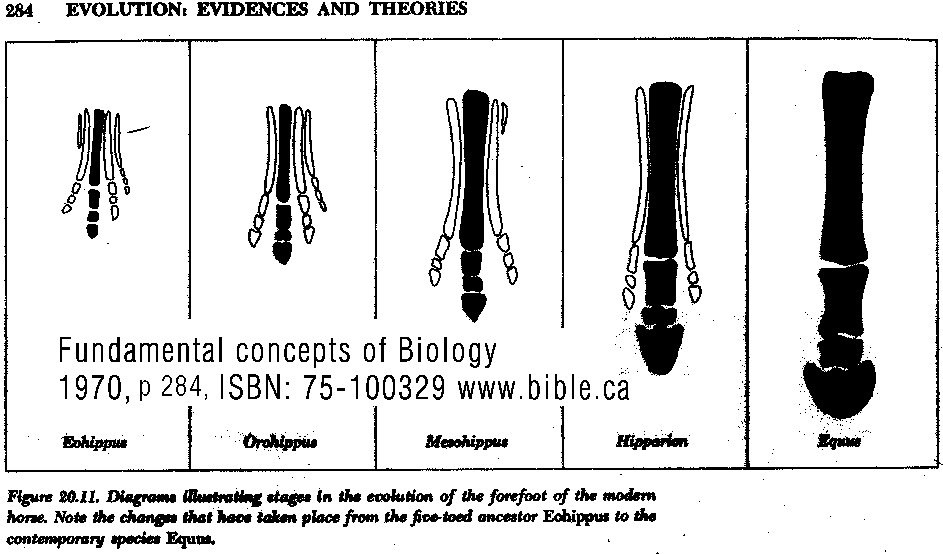
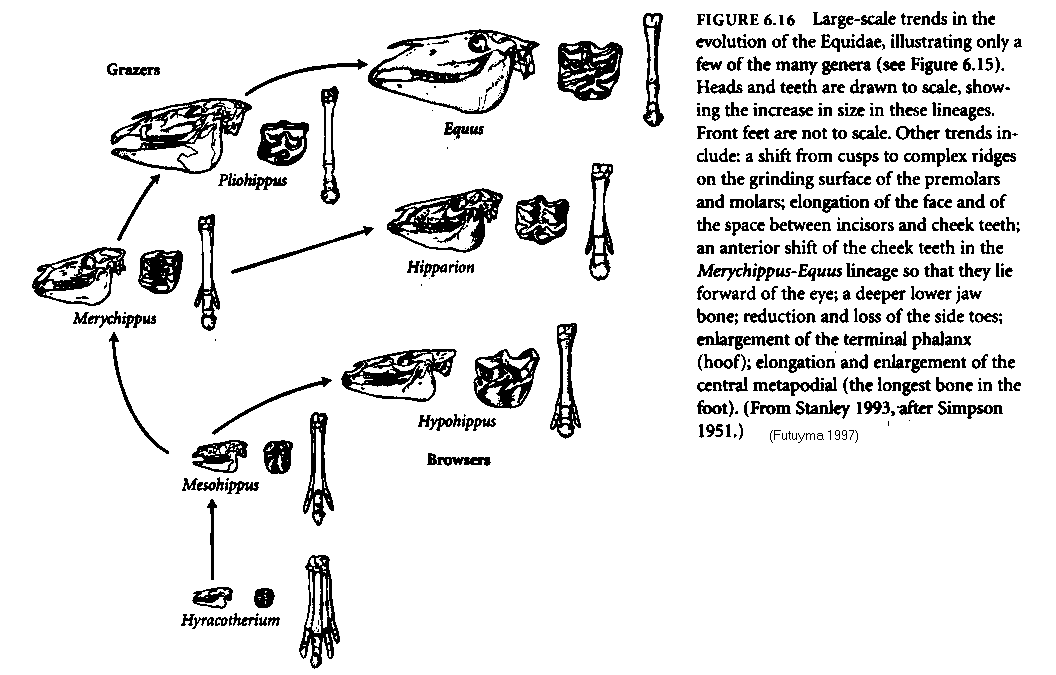
Although single characters do not convey an adequate sense of the evolution of the organism as a whole, they are easier to describe in quantitative terms. Because of the mosaic nature of evolution, this approach often suffices to describe the rate of evolution of taxonomically or adaptively important features. Thus one may describe the change in, say, the height of a horse's tooth in millimeters per million years. Evolutionary rates are usually described in relative terms. Two measures are often used (Box 6.A). The number of standard deviations by which the mean of the character changes per unit of time scales the change to the amount of variation within the population (Figure 6.36); thus if the mean changes by three standard deviations, it has shifted to a phenotypic value that formerly characterized less than 1 percent of the population. Another measure of evolutionary rate is the darwin, which is a change by a factor of 2.718 (the base of natural logarithms) per million years. The paleontological literature on rates of evolution does not necessarily represent a random sample of evolutionary rates, because paleontologists often focus on characters that display evolutionary change and ignore characters that do not distinguish taxa (i.e., CONSERVATIVE CHARACTERS, those with a very low rate of evolution). When rates of character evolution have been measured for ancestor-descendant series of dated fossils, the most striking result is that average rates of evolution are usually extremely low. For example, Figure 6.37 shows the rate of evolution of the height of one of the molar teeth in a lineage of horses, one of the most important adaptive changes in the evolution of the grazing lineages. The maximum rate was 94 millidarwins (about 7 percent per million years). Expressed another way, the maximum rate was 1.7 standard deviations per million years, and the average rate was 2.8 standard deviations per million generations, meaning that it took this long for the average tooth height to shift less than one-third of the span between the smallest and the largest specimens that the population harbored at any one time. Another example, again from the Equidae, is provided in Figure 6.38, which presents the estimated body weights of fossil equids and the rate of change in body weight for some ancestor-descendant series. Note that the rate of evolution of this feature was very low until the early Miocene (25-15 Mya), when the horses began an adaptive radiation into diverse browsing and grazing lineages, some of which increased in size rather rapidly. Even the most rapid rates are quite low, however: 280 millidarwins, or about a 21 per-cent increase in size per million years on a logarithmic scale. (Note also that some lineages decreased in size, at various rates.) These are average rates, calculated from the difference between the mean values at the beginning and at the end of a time interval, divided by the interval [(Y1, - Y0)/t]. In all cases the time interval is on the order of millions of years. One can readily see that very rapid evolutionary rates would be masked if within this interval the rate fluctuates (e.g., long periods of stasis alternate with bursts of very rapid change) or the direction of evolution changes (e.g., the mean increases and then declines to its original value, yielding no net change). Such changes in rate and direction are indeed evident when successive fossil deposits are close-ly spaced in time, as the data from sticklebacks and plank-tonic protozoans illustrate (see Figures 6.6-6.8). In almost all instances in which data of this kind have been analyzed (Charlesworth 1984), the variation in the rate of change between successive samples is much greater than the average rate of change over the whole sequence, largely because of rapid fluctuations in the mean. Over very short intervals, rates of change may be very rapid, in the tens or even hundreds of darwins (Figure 6.39). Differences between Pleistocene species (less than a million years old) and their living descendants represent some very rapid rates of change, and even faster changes have been documented in populations that have been introduced by humans into new geographic regions. For ex- ample, house sparrows (Passer domesticus), introduced into North America from Europe about 100 years ago, have be- come differentiated into geographic races that are adapted in size and coloration to different North American environments (Johnston and Selander 1,964). Some of their skeletal dimensions have diverged from those of European populations at rates of 50 to 300 darwins. In absolute terms, many of the changes in introduced or post- Pleistocene populations have been slight, and would not be noticed without measuring them; but they have transpired over such a short time that the rate of change is very great. The overall messages of the data in Figure 6.39 are that in discussing evolutionary rates, it is important to specify the time interval involved, and that a high rate of evolution is not sustained in one direction for very long. Both proponents and opponents of the hypothesis of punctuated equilibrium have claimed that data on evolutionary rates support their views. "Punctuationists" expect the average evolutionary rate to be low, because they expect brief intervals of rapid change to punctuate much longer periods of stasis. The Globorotalia lineage (see Figure 6.6), in which there was a rapid shift from one relatively stable phenotype to another, is an example of the pattern they predict. Opponents of punctuated equilibrium hold that the rapid fluctuations in these characters, even when there is no overall directional trend, provide strong evidence against punctuated equilibrium. These features, they argue, are not constrained from evolving, as the punctuationists propose; rather, the characters are continually evolving, but simply are not going in any particular direction. (Evolutionary Biology, Douglas J. Futuyma, Third Edition, 1998 ISBN: 0-87893-189-9, p 159-161)

(Click on photo for high resolution)
FIGURE 6.37 [above left] Analysis of the rate of evolution of a morphological character in the fossil record. Changes in the height of a molar tooth in the horse lineage Hyracotherium to Neohipparion are plotted arithmetically (black circles) and logarithmically (colored circles). The average standard deviation (s = 0.055) on the log scale is shown by horizontal bars. For each interval in this sequence, several calculations are shown: (1) the estimated number of generations (g) between successive time points; (2) the change in mean tooth height (A P), measured in standard deviations on the log scale; and (3) the average fraction (f) of the population dying, per generation, if natural selection were responsible for the change and if the change occurred at a constant rate during the time interval. Over this history, the average rate of change was 2.8 mm per million generations (on an arithmetic scale), or 2.6 standard deviations per million generations (on a log scale). The average fraction of mortality that would account for this rate of change, if natural selection were responsible, is 1.0 x 10--6 of the population per generation (see Chapter 14). (After analyses by Lande 1976a.) (Evolutionary Biology, Douglas J. Futuyma, Third Edition, 1998 ISBN: 0-87893-189-9, p 161)
FIGURE 6.38 [above right] Evolution of estimated body mass in fossil horses. (A) The body masses of 40 species, plotted against geological time. Notice that although the mean increases toward the present (an instance of Cope's rule), this is largely because of an increase in variation; small species existed throughout much of the history of the Equidae. (B) Rates of evolution of body mass (in darwins) between various pairs of ancestral and descendant taxa, plotted against the midpoint between their geological ages. Both increases (solid circles) and decreases (open circles) in size occurred. The most rapid changes occurred late in equid history, during the evolution of large grazers. Even the highest rate of evolution is much lower than some rates calculated over shorter intervals (see text). (After MacFadden 1986.) (Evolutionary Biology, Douglas J. Futuyma, Third Edition, 1998 ISBN: 0878931899, p 161) |
|
Biology in Action, N. J. Berrill, Strathcoma Professor of Zoology, McGill University, p 790 |
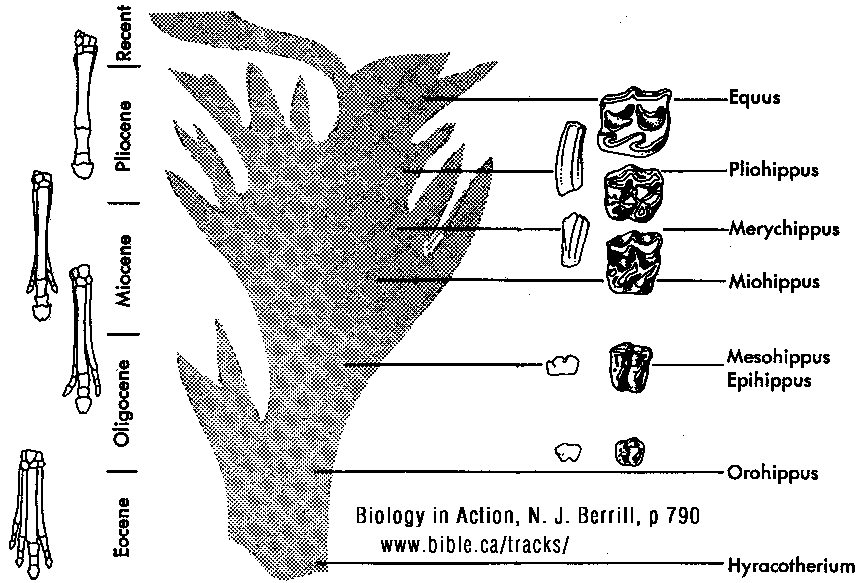
(Click on photo to see high resolution)
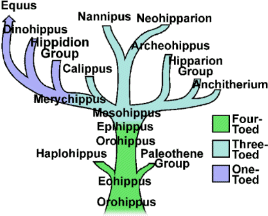
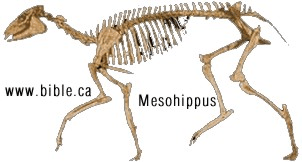
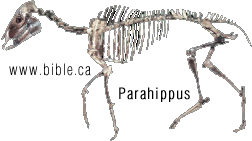
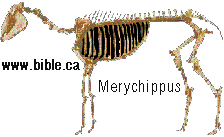
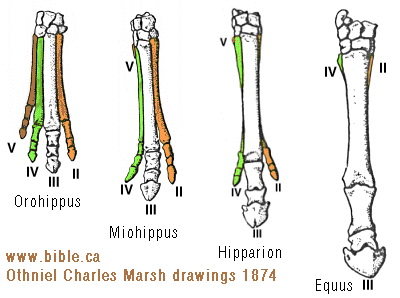
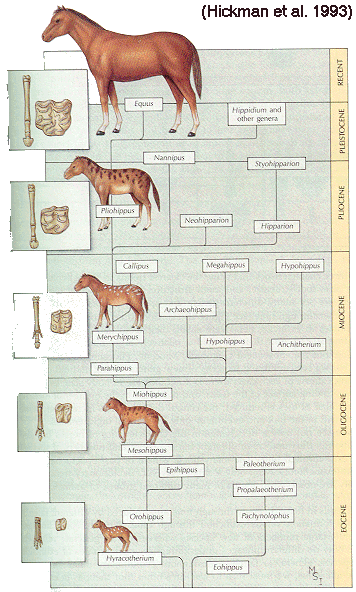
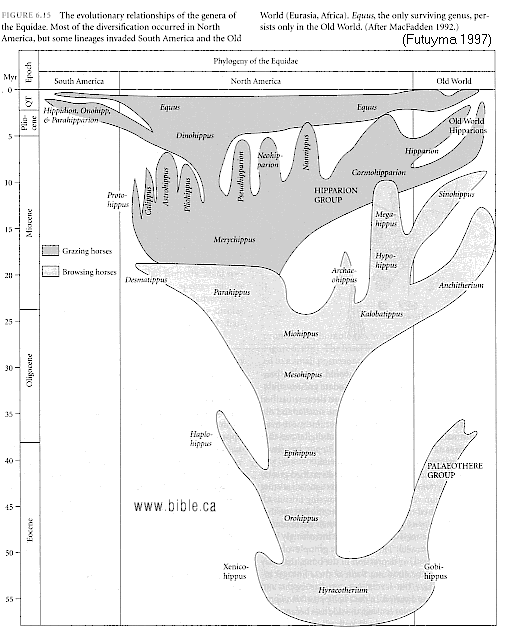
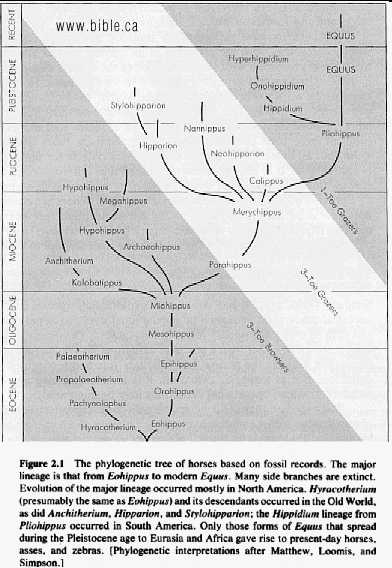
HORSE EVOLUTION: When an attempt is made to trace back the evolutionary history of any group of animals or plants we inevitably find ourselves groping much further into the past than was anticipated. To find the origins of the phyla themselves, the basic divisions, we cross the Cambrian period into Pre-Cambrian time and lose the trails. To find the beginnings of life on land the indications, for both animal and plant, lead us as far back as the Silurian. To find the first signs of reptilian life we must go back to the Carboniferous, with its predominant coal-forest-amphibian assembly. To find mammalian trends among the reptiles we go back as far as the early Permian, long before the reptiles them-selves had evolved to any great extent. Birds and flowering trees were opening their door to the future in time to see the Age of Reptiles get under way, and in like manner the mammals were already fully mammalian, widespread, and diversifying during the Cretaceous, well be-fore the Age of Mammals was worthy of its name. Each order and most of the families of mammals seem to have become set in their courses at least from the very beginning of the Cenozoic Age, in each case pointing in a certain general direction and, like a rocket shot through time, diverging more and more from its companions; and all the time changing into diverse forms during its passage, some to succeed but most to falter and fail. Two cases signify more than most: the horse family, the Equidae, because its evolutionary history is so completely documented by fossils, and the order Primates, because we ourselves are its most successful survivors. The evolution of horses is a progressive adaptation of originally unspecialized mammals to life on the plains, where grasses are the principal food and predators a constant menace. Primate history has been mainly progressive adaptation to arboreal life above the ground, a relatively safe but equally specialized way of life. In each case once the initial trend became established, the general course of evolution appears inevitable in retrospect. Horse Evolution: We have already referred to the evolution of horses in connection with the rate of evolutionary change and in an incidental way in the general account of the Cenozoic era. The evolution of the family is given here in some-what greater detail to illustrate further the pattern of survival, the force of selection, and also to supply a sort of comparative calendar of events against which the evolution of the primates, including man, can be more clearly seen. The first members of the horse family became recognizable early in the Eocene fifty or so millions of years ago, in the form of Hyracotherium (better known as Eohippus, a name which means dawn horse). Hyracotherium did not look in any way like a horse, being about the size of a cat, with a flexible back and a long Sail. Its face was short and its eyes near the front, but its toes were equipped with hoofs, with the little toe of the front feet and both the little and big toes 'of the hind feet already beginning to dwindle. In other words, its feet were already designed for speed and a running start. And even in Hyracotherium the teeth show the first signs of specialization characteristic of horses, with distinct cusps and a trend toward strengthening and enduring. TEETH AND SURVIVAL The evolution of teeth has played a great role not only in the evolution of horses, and in fact in the evolution of vertebrates as a whole, but because of their hardness and consequent survival as fossils, teeth have often been the main key to deciphering the fossil record (Fig. 38-1). So far as the animal itself is concerned the teeth have a double significance. An animal can feed and live only until its teeth wear out. Fish and reptiles have no problem in this respect since replacement of old teeth by new continues indefinitely. Birds bypass the issue by having no teeth at all and are able to maintain the horny beak with ease. Mammals have reduced their tooth replacement to a single occasion the replacement of the milk teeth of the small jaws of the very young by the one and only set of permanent teeth. With the exception of man, when these teeth wear out, the animal dies of starvation no matter how healthy its body may have been until then. As a general rule larger animals live much longer than small ones. Their teeth must there-fore be correspondingly large to serve their immediate function and must also endure for periods corresponding to the potential life span of the animal. With the exception of elephants, even the largest mammals become senescent and die. in their third decade, and at the end of twenty years or so of incessant use, most mammalian teeth have had their day. Elephants, which may live four times as long and must continue to eat, have huge complex molars for grinding heavy vegetation. Even an elephant molar wears out long before an elephant does, and the problem has been solved by having the several molars of each half move into position and action in succession instead of all at once, thereby extending the potential life of the animal to over 70 years. Mammalian teeth are distinctive in being differentiated. Reptilian teeth are essentially all alike, but even in the most primitive mammals, the insectivores, the teeth of each half -jaw are differentiated into three incisors, one canine, four premolars, and three molars, a dental formula of 3.1.4.3/ 3.1.4.3 for upper and lower half-jaws. Man has departed from this basic mammalian dental formula to only a moderate extent, having lost one incisor and two pre-molars in each set. The human dental formula is 2.1.2.3/2.1.2.3 a formula which we have in common with the Old World monkeys and apes. In horses and grazing mammals generally, the canines and first premolars are lost, with the emphasis placed on incisors for cropping grass and molars and the more posterior premolars for grinding this tough siliceous material. A large part of horse evolution has been as much an adaptation of the animal to certain general changes in its own nature as it has to external circumstance. Because of the incessant threat of increasingly fast, large, and efficient predators, the horse family for the most part has been relentlessly driven toward becoming larger and, if possible, faster. Whether the modern horse can run any faster than could Hyracotherium is a moot point. It is significant that a racing horse and the small, slender racing greyhound dog of today have much the same top speed, about 35 miles per hour. Hyracotherium, with its arched, flexible back like that of a greyhound, hoofs for protecting hard-driven feet from the wear and tear of impact against the ground, and reduced lateral toes indicating that it ran on its central toes only, was as much built for speed as any animal has ever been. Its need for speed was to escape the carnivorous catlike predators of the time. Increase in body size would have lessened vulnerability with regard to the small predators, but as predatory animals gradually increased in size (concurrently with the appearance of larger prey), the advantage would be lost and further increase in size would be desirable. Progressive increase in size meant not only increasing weight to be propelled over the ground but also a gradual lengthening of the span of life to several times its original duration. Much of the change that we see in the evolution of the modern horse from its small Eocene ancestor has been a progressive adjustment of structure to the demands of increasing mass and time, assuming that the initial efficiency at least must be maintained. Above all, legs and muscles must propel forward an ever -increasing mass without loss of speed or the wearing out of feet, and teeth must last longer and longer as the natural life span becomes extended. Increasing body height involved a lengthening of the neck and head so that the mouth could be used for close-cropping grass. The forepart of the face lengthened; the eyes were kept high and were shifted to the side to raise vision above grass level to watch for predators. Leg and foot bones enlarged, lengthened, and fused along the weight- supporting axis, at the expense of lateral components, including toes. Operating muscles became short and bulky and associated only with the upper parts of the limbs. Hoofs have been no problem, once in existence, since the horny hoof, evolved from a horny claw, is continually growing through-out an animal's life, just as do our own nails. Teeth became hardened with cement to cope with the siliceous character of grasses, and lengthened so as to serve the longer life. None of these changes imply any increase in efficiency but only the maintenance of original efficiency of the small, fast, and relatively short-lived primitive creature represented by Hyracotherium. HORSES AND PLAINS Disregarding the curtain raiser in which typically five-toed herbivorous Paleocene mammals, of a kind now extinct, gave rise to the first small members of the hoofed, odd-toed members of the horse family, the first scene opens in North America in the early Eocene. At this time the land bridge connecting North America and Asia, at the site of the Bering Strait, was in existence and Hyracotherium spread across into Asia, where it persisted for a while, then died out. By the middle Eocene the land connection was replaced by a water barrier, and horse evolution continued mainly in North America through the remainder of the Eocene and through the Oligocene epochs. During this period the Hyracotherium stock grew to sheep size and flourished as the genus Miohippus, a slender type with an arched back, three toes now on both fore and hind feet, and the median bone in each foot becoming a straight, cylindrical cannon bone. The first three-toed horses now existed. They had a better brain, eyes farther to the side, and ran more on their. toes. Premolar as well as molar teeth became specialized for grinding the abrasive grass. At the same time both the true cats and the saber-toothed cats were larger and more powerful. The second scene begins with the Miocene and is associated with a cooling climate, elevating lands, restriction of forests, and expansion of grasslands. Certain of the primitive horses took more to the forests and survived as somewhat flat-footed forest horses for mil-lions of years, some of them crossing a temporarily re-established bridge to Asia, only to die out eventually. The main trend was toward larger size, longer heads with eyes set farther back, more powerfully constructed legs for running, and side toes that now barely touched the ground. Teeth became longer and harder. Such was the nature of Parahippus, the most successful of the early Miocene horses, which gradually gave way to more progressive, rapidly evolving descendants called Merychippus. These had long grazing teeth well embedded in the bony jaws for greater anchorage. The herds of Merychippus far outnumbered all others. Then in the Pliocene came the greatest surge of horse evolution, associated with further expansion of grassland plains and steppes. This was, in fact, a period of great expansion for other groups of large mammals, particularly the family of elephants which were to give rise to the elephants, mammoths, and various kinds of mastodons of later times. The land bridge between Asia and North America again became established during this time, and the three-toed horses spread across into Asia from their North American homeland. These invading horses, descended from Merychippus and known as Hipparion, flourished in Eurasia for some millions of years, giving rise to several new kinds, although Hipparion itself had died out in North America. Other descendants of Merychippus in North America, however, gave rise to several other types, such as the medium-sized, slender, three-toed Nannihippus, which in turn yielded the larger and equally abundant three-toed Neohipparion, the two coexisting for about 11 million years. Hipparion in Asia and the other two pre-horses in North America survived abundantly until the end of the Pliocene, when the. Pleistocene Ice Age was already imminent. In North America Merychippus gave rise to a group of larger, one-toed horses known as Pliohippus, and it is from this animal that the presently surviving horses-the horse proper, the ass, and the zebra-arose. All the other stocks, including some not mentioned in the foregoing account, died out at the end of the Pliocene, or in the early Pleistocene, with the increasing onslaught of the cold from the north. With the Ice Age imminent, the history of the horse family continues in a single narrow line, Equus, the true horses, representing a single branch of the wide spreading family tree of the past 50 million years. At every level of general advance, one branch usually has been exceptionally successful and has given rise to branches of its own, while other, older branches have come to the ending of their line. In the early Pleistocene the true horses, including the zebras and asses, had spread not only over North America but into Eurasia-the third great dispersal into that area-where they displaced Hipparion. (Biology in Action, N. J. Berrill, Strathcoma Professor of Zoology, McGill University, p 790) |