School Textbook Fraud:
The Pepper Moth Theory Falsified

|
School Textbook Fraud: The Pepper Moth Theory Falsified |
|
"Piltdown Moth" by Steve Rudd
![]() see also "Mothballed Science" by Phillip E. Johnson
see also "Mothballed Science" by Phillip E. Johnson
|
|
|
|
Textbook Fraud
|
Biology Textbook Fraud
|
|
|
|
|
The Pepper Moth:
Click on photos for high resolution
|
What was observed: |
|
|
|
|
Another way of looking at the Pepper moth story A simple analogy |
If the two varieties of moth (the typica or white variety and the carbonaria or black variety) were say, black and white soldiers on a battlefield at night, it would be reasonable to say that the white soldiers would have more casualties. Eventually, only black soldiers would remain but no one would claim this to be evolution in action! Recently there have been reports from Uganda that about 15% of the elephants are now born without tusks. This is the result of a mutation which prevents tusks from developing, but selection is the logical explanation. Like the white soldiers in the above example, ivory poachers kill only those elephants with a fine set of tusks leaving the tuskless type to multiply. It is quite illogical to call industrial melanism in the story of the black and white moths or the black and white soldiers or the tuskless elephants evidence of evolution. These are examples of natural selection but no new genetic information has been added, no new creature has been created. (Comments by Ian Taylor, who is also the author of treasure, "In the Minds of Men") |
|
|
|
Epicycles and The Pepper Moth Theory!
|
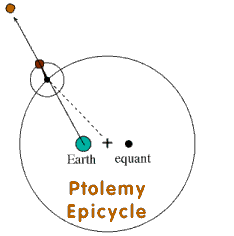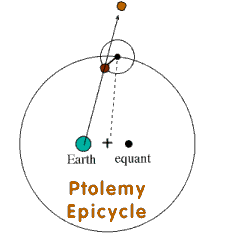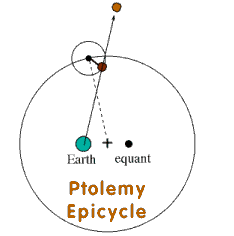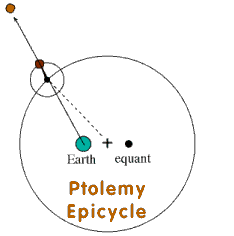
Ptolemy's Epicycles
Ptolemy started with the wrong theory that the earth was the center of the universe and the sun and planets rotated around the earth. In order to explain the unusual motion of the planets, he came up with the machine that used what he called "epicycles". Using combinations of opposite and concurring epicycles, Ptolemy was able to create a model of the universe that explained the actual observed motions of the planets with very high accuracy. For example Mars needed two epicycles and three to explain the motion of Jupiter. But as well as the theory fit the data, it was wrong. The Pepper moth is exactly like this. |
Pepper Moth Theory Epicycles
The Pepper moth theory is identical to the Ptolemy epicycle system. Sun = Pepper moth The observed data of melanics in moths is founded upon the false notion of selective bird predation as the primary force. The pepper moth theory corresponds to the Sun in the Ptolemy system and the theory fits the data easily. However in order to harmonize the vast majority other moths (other planets) that are also melanic where bird predation and industrial pollution are not a factor at all, Majerus and other evolutionists must invent new theories (epicycles) to explain the otherwise unexplainable. When even epicycles won't help, the magic wand of "mutations" is used to explain the rest. The result is a basic theory with so many modifications (epicycle upon epicycle upon epicycle like Jupiter), that it is simply too complicated to be true. Is Majerus the echo spirit of Ptolemy? The variation seen in the Pepper moth, is quite common in the natural world. For example white tail and mule deer are two breeds of the same species, and totally cross fertile. However they have slightly different coloration, quite different antlers and live in distinct habitats. Bird predation has about as much to do with the variation of melanics in Pepper moths, as it does in deer. Selective predation by birds upon the Pepper moth must be rejected along side of the false notion that the sun revolves around the earth. Evolutionists, like Ptolemy both took their theory down a dead end road, both are false. White Grizzlies (polar bears) life in the Arctic and dark Grizzlies live in the rocky mountains. Selective bird predation cannot explain moths, deer or bears etc. Evolutionists can no more explain the mechanism that produces the observed variation in bears and deer than they can the Pepper Moth. Creation, on the other hand, unlocks the key to understanding the pepper moth because God simply created moths, deer, dogs, cats, bears etc in the beginning with the genetic variation needed to produce what we are presently observing "against the stars". Like Ptolemy, Majerus' pepper moth theories do explain what we see, but both are wrong. |

|
|
Use this fraud test on your own text books: We charge any school textbook with fraud and gross misrepresentation if it:
|
|
|
|
|
Why the Pepper Moth is so important to evolutionists (What evolutionists say about it) |
|
Informed scientists reject the Pepper Moth Theory as taught in School Textbooks |
|
|
|
|
|
The Pepper Moth Fraud Exposed Important Summary |
|
|
|
|
|
The Pepper Moth theory is falsified because:
|
|
|
|
|
Components of the 'textbook' peppered moth story Falsified

|
|
Why it is falsified |
|
(a) The peppered moth has two distinct forms, one, the typical form, being white with black speckling, the other, I. carbonaria, being almost completely black. |
False! There are not two forms, but hundreds. There are light and dark and every shade inbetween. "If a large number of peppered moths, collected across the British Isles, are examined, it is obvious that the majority of individuals are either f. typica or f. carbonaria. However, a significant proportion of individuals do not fit the descriptions of either of these forms. The variation in patterning extends from white, with just a very fine dusting of dark scales, to almost completely black, with all grades of intermediates between the two (Bowater 1914)." (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p117) |
|
(b) Peppered moths fly at night and rest on tree trunks by day. |
The fact is, that Kettlewell had to glue two moths, a black one and a white one, on a tree trunk to take the classic photograph that has appeared in every school textbook for over forty years! Peppermoths are nocturnal, but do not rest on tree trunks. We are 100% certain that pepper moths do not rest on tree trunks. Pepper moths rest during the day high up in trees in unexposed dark shaded areas under lateral branches in the leaves. |
|
(c) Birds find peppered moths on tree trunks and eat them. |
Birds do eat moths that rest on tree trunks, but the pepper moth does not rest on tree trunks. In fact birds may not find any Pepper Moths where they actually hide high up in the twigs and leaves. |
|
(d) The ease with which birds find peppered moths on tree trunks depends on how well the peppered moths are camouflaged. |
Why did birds eliminate in polluted forests, the light coloured pepper moths that rest high in trees hidden under branches, but DID NOT eliminate the light coloured Oak Beauty moths that do rest on the dark tree trunks? "Many nocturnal moths do rest by day on tree trunks and searching trunks has long been a recognised method of collecting employed by lepidopterists. However, the number of published records of peppered moths being found on tree trunks is negligible." (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p121) |
|
(e) Typical peppered moths are better camouflaged than melanics on lichen-covered tree trunks in unpolluted regions, while melanics are better camouflaged in industrial regions where tree trunks have been denuded of lichens and blackened by atmospheric pollution. |
To humans, a typical pepper moth on foliose lichens appears camouflaged, however, since birds see in UV, these same Pepper Moths stand out like a white moth on a black tree trunk. This one fact alone completely falsifies the Pepper Moth theory. Jim Stalker, has assessed the UV reflectance characteristics of all the common forms of the peppered moth, together with six lichen taxa taken from seven tree species. He found that while the black scales and hairs of all forms of the peppered moth absorb UV wavelengths, the white scales reflect UV strongly. Stalker's results in respect of lichens are of particular interest. A number of authors have stressed the correlation between the frequency of typica and foliose or vegetative lichens. Stalker thus considered foliose and crustose lichens separately. All parts of the foliose lichens analysed, including Hypogymnia and Evernia spp., absorbed UV. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p136) Pepper Moths do not rest on tree trucks, so the change in the colour of the tree trunks cannot be a factor in making dark moths more camouflaged. |
Pepper Moth Story does not belong in School Textbooks!
|
|
|
|
Uncertain Things about the Pepper Moth theory that need more study: Michael E. N. Majerus |
|
|
|
|
|
|
|
||
|
Photo Gallery of the Pepper Moth |
|||
|
|
|
||
|
|
The Pepper Moth: Dark coloured Melanic (biston betularia carbonaria) |
||
|
|
|
||
|
|
The Pepper Moth: Light coloured typica (biston betularia typica) |
||
|
|
|
||
|
Photo copyright by David Painter |
Oak Beauty (Biston strataria)A trunk rester that is the closest relative to the Pepper moth and have very similar habits, except the Pepper moth is not a trunk rester. Both the pepper moth and the Oak Beauty moth, have a dark (melanic) and light (typica) variation. Yet the proportion of light and dark Oak Beauty moths did not experience a large increase in melanics as is reported for the pepper moth. This in spite of the fact that the Oak Beauty moth normally rests on tree trunks and the pepper moth does not. This fact alone completely falsifies the idea that bird predation is responsible for eliminating light coloured pepper moths in dark polluted forests. Why did birds eliminate in polluted forests, the light coloured pepper moths that rest high in trees hidden under branches, but DID NOT eliminate the light coloured Oak Beauty moths that do rest on the dark tree trunks. The same tree has typica (light) pepper moths resting hidden high in its branches and typica (light) Oak Beauty moths resting below in plain view on the trunk, yet the bird eliminated the hidden ones, but left the ones in plain view? Not likely. |
||
|
|
|
||
|
Brindled Beauty (lycia hirtaria)A trunk rester that like the Oak Beauty, never experienced more than a very slight increase in the frequency of the melanic form of the moth. Why did "selective bird predation" have almost no effect upon the "Brindled Beauty? Photo copyright by David Painter |
|||
|
|
|
||
|
|
Pale Brindled Beauty (Apocheima pilosaria)Why did "selective bird predation" have almost no effect upon the "Pale Brindled Beauty? The melanic form never outnumbered the light coloured typical by almost 4 times. Photo copyright by Ian Kimber |
||
|
|
|
||
|
The Diurnea Fagella (melanic form) unlike the Pepper Moth, is a trunk rester that falsifies the Pepper moth theory. Although eaten by birds, the frequency of melanic and typical moths does not correspond to industrial areas as the Pepper moth is alleged to conform to because of selective bird predation. So here we have a known trunk resting moth that selective bird predation has no effect upon, whereas the Pepper moth doesn't even rest on tree trunks and likely hides high in the canopy hidden under leaves where birds rarely find them during the day. |
|||
|
|
|
||
|
Photo copyright by Ian Kimber |
Buff Ermine (spilosoma luteum)Here we have a moth, the Buff Ermine, that is not eaten by birds because of it toxicity. Obviously selective predation by birds was not a factor in this case of melanism. Although Majerus admits the moth is unstudied, he ventures to guess that the moth evolved into the melanic form in order to not be detected by predators. But if the moth is not eaten by birds, what predators are we talking about? He sort of makes it appear that the moth had to sit down and weigh the benefits of such a change then decide to trade off its aposematic coloration for camouflage. However such a change is not even needed because the typical moth survives just fine! Evolution theory needs many epicycles to make it work! |
|
|
|
||
|
click on photo for high resolution |
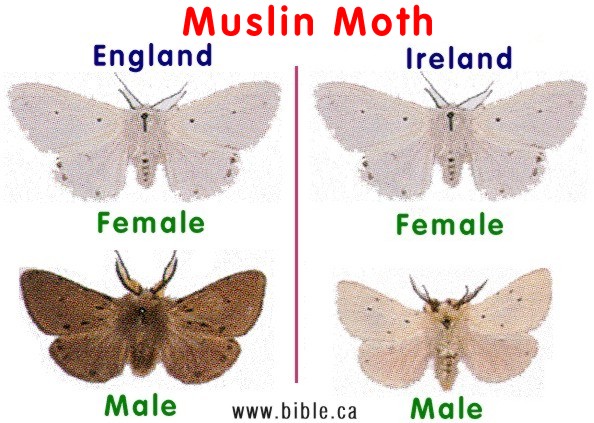
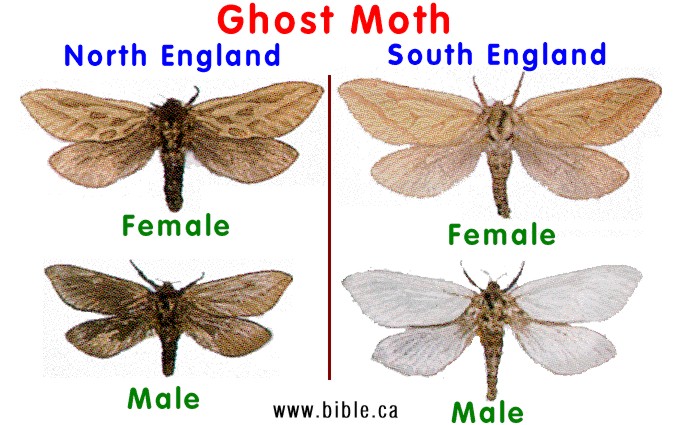
The Muslin moth:If birds will not eat this moth because they are "distasteful", then selective bird predation is not a factor to explain why males are melanic in England and white in Ireland. The Muslin moth has dark males and white females, whereas the Ghost moth has white males and dark females in the same area. When we compare four moths, the Muslin, Ghost, Oak Beauty and Pepper moths, it becomes quite obvious that bird predation is not a cause for melanics in any of these species. click for more |
||
|
|
|
||
|
click on photo for high resolution |
The Ghost moth |
||
|
|
|
Book Review of Majerus' book on Melanism
|
|
|
|
Jerry A. Coyne's review of Majerus' book on Melanism Jerry A. Coyne is in the Department of Ecology and Evolution, University of Chicago, 1101 E. 57 Street, Chicago, Illinois 60637, USA. (Nature 396, 35 - 36 (1998) © Macmillan Publishers Ltd.)
|
"Not black and white" From time to time, evolutionists re-examine a classic experimental study and find, to their horror, that it is flawed or downright wrong. We no longer use chromosomal polymorphism in Drosophila pseudoobscura to demonstrate heterozygous advantage, flower-colour variation in Linanthus parryae to illustrate random genetic drift, or the viceroy and monarch butterflies to exemplify Batesian mimicry. Until now, however, the prize horse in our stable of examples has been the evolution of 'industrial melanism' in the peppered moth, Biston betularia, presented by most teachers and textbooks as the paradigm of natural selection and evolution occurring within a human lifetime. The re-examination of this tale is the centrepiece of Michael Majerus's book, Melanism: Evolution in Action. Depressingly, Majerus shows that this classic example is in bad shape, and, while not yet ready for the glue factory, needs serious attention. According to the standard textbook litany, before the mid-nineteenth century, all B. betularia in England were white moths peppered with black spots, a form called typica. Between 1850 and 1920, typica was largely replaced by a pure black form (carbonaria) produced by a single dominant allele, the frequency of which rose to nearly 100% in some areas. After 1950, this trend reversed, making carbonaria rare and typica again common. These persistent and directional changes implied natural selection. In a series of studies, this conclusion was verified by several investigators, most prominently Bernard Kettlewell of Oxford. According to these workers, the evolution of colour was caused by birds eating the moths most conspicuous on their normal resting site -- tree trunks. The increase in black moths was attributed to pollution accompanying the rise of heavy industry. A combination of soot and acid rain darkened trees by first killing the lichens that festooned them and then blackening the naked trunks. The typica form, previously camouflaged on lichens, thus became conspicuous and heavily predated, while the less visible carbonaria enjoyed protection and increased in frequency. After the passage of the Clean Air Acts in the 1950s, trees regained their former appearance, reversing the selective advantage of the morphs. This conclusion was bolstered by a geographical correlation between pollution levels and morph frequencies (carbonaria was most common in industrial areas), and most prominently by Kettlewell's famous experiments which showed that, after releasing typica and carbonaria in both polluted and unpolluted woods, researchers recaptured many more of the cryptic than of the conspicuous form. The differential predation was supported by direct observation of birds eating moths placed on trees. Finally, Kettlewell demonstrated in the laboratory that each form had a behavioural preference to settle on backgrounds that matched its colour. Criticisms of this story have circulated in samizdat for several years, but Majerus summarizes them for the first time in print in an absorbing two-chapter critique (coincidentally, a similar analysis [Sargent et al., Evol. Biol. 30, 299-322; 1998] has just appeared). Majerus notes that the most serious problem is that B. betularia probably does not rest on tree trunks -- exactly two moths have been seen in such a position in more than 40 years of intensive search. The natural resting spots are, in fact, a mystery. This alone invalidates Kettlewell's release-recapture experiments, as moths were released by placing them directly onto tree trunks, where they are highly visible to bird predators. (Kettlewell also released his moths during the day, while they normally choose resting places at night.) The story is further eroded by noting that the resurgence of typica occurred well before lichens recolonized the polluted trees, and that a parallel increase and decrease of the melanic form also occurred in industrial areas of the United States, where there was no change in the abundance of the lichens that supposedly play such an important role. Finally, the results of Kettlewell's behavioural experiments were not replicated in later studies: moths have no tendency to choose matching backgrounds. Majerus finds many other flaws in the work, but they are too numerous to list here. I unearthed additional problems when, embarrassed at having taught the standard Biston story for years, I read Kettlewell's papers for the first time. Majerus concludes, reasonably, that all we can deduce from this story is that it is a case of rapid evolution, probably involving pollution and bird predation. I would, however, replace "probably" with "perhaps". B. betularia shows the footprint of natural selection, but we have not yet seen the feet. Majerus finds some solace in his analysis, claiming that the true story is likely to be more complex and therefore more interesting, but one senses that he is making a virtue of necessity. My own reaction resembles the dismay attending my discovery, at the age of six, that it was my father and not Santa who brought the presents on Christmas Eve. Occupying a quarter of the book, the Biston analysis is necessary reading for all evolutionists, as are the introductory chapters on the nature of melanism, its distribution among animals, and its proposed causes. Majerus, however, designed his book for both professional and lay readers, and this causes some unevenness in the material. The Biston story is sandwiched between less compelling chapters, including long sections on the basic principles of genetics and evolution, which can be skipped by evolutionists. Other discussions, involving melanism in ladybirds and other Lepidoptera, as well as the author's unpublished work on habitat selection, are full of technical details that will overwhelm the lay reader. Unfortunately, most of the work described is inconclusive; despite the widespread occurrence of melanism, its evolutionary significance is nearly always unknown. What can one make of all this? Majerus concludes with the usual call for more research, but several lessons are already at hand. First, for the time being we must discard Biston as a well-understood example of natural selection in action, although it is clearly a case of evolution. There are many studies more appropriate for use in the classroom, including the classic work of Peter and Rosemary Grant on beak-size evolution in Galapagos finches. It is also worth pondering why there has been general and unquestioned acceptance of Kettlewell's work. Perhaps such powerful stories discourage close scrutiny. Moreover, in evolutionary biology there is little payoff in repeating other people's experiments, and, unlike molecular biology, our field is not self-correcting because few studies depend on the accuracy of earlier ones. Finally, teachers such as myself often neglect original papers in favour of shorter textbook summaries, which bleach the blemishes from complicated experiments. It is clear that, as with most other work in evolutionary biology, understanding selection in Biston will require much more information about the animal's habits. Evolutionists may bridle at such a conclusion, because ecological data are very hard to gather. Nevertheless, there is no other way to unravel the forces changing a character. We must stop pretending that we understand the course of natural selection as soon as we have calculated the relative fitness of different traits. |
|
|
|
|
|
|
|||
|
|
"Second Thoughts about Peppered Moths" Exerted from the chapter: "Problems with the classic story" Researched and written by Jonathan Wells, Ph.D., April 1999 |
|||
|
|
|
|||
|
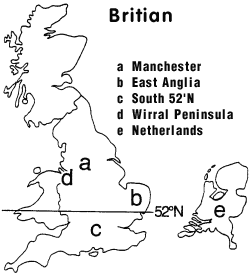
Problems with Pepper Moth Theory: The U.K. and The Netherlands. |
U.K. |
|||
|
Map Key: |
|
|||
|
This never happened, however, indicating that factors other than selective predation must be affecting melanic frequencies. Some investigators attributed the discrepancy to heterozygote advantage (Clarke and Sheppard 1966, Lees and Creed 1975), but it has since been established that there is no evidence for this (Creed et al. 1980, Lees 1981, Mani 1982, Cook et al. 1986). (b) The proportion of melanics in East Anglia reached 80% despite the absence of any apparent pollution (Lees and Creed 1975); after the introduction of pollution control legislation, typicals became predominant before lichens returned to the trees (Grant and Howlett 1988). ... Some other distribution features were inconsistent with Kettlewell's explanation, as well. In rural Wales, the frequency of melanics was higher than expected, leading Bishop to conclude that "as yet unknown factors" were involved (Bishop 1972, p. 240). In rural East Anglia, where there was little industrial pollution and typicals seemed better camouflaged, melanics reached a frequency of 80%, prompting Lees and Creed to write: "We conclude therefore that either the predation experiments and tests of conspicuousness to humans are misleading, or some factors or factors in addition to selective predation are responsible for maintaining the high melanic frequencies" (Lees and Creed 1975, pp. 75-76).(c) South of latitude 52*N, the relatively poor correlation of melanism with sulfur dioxide concentration suggested that non-industrial factors were of greater importance than selective predation (Steward 1977a,b); after the introduction of pollution control legislation, the proportion of melanics decreased in the north, as expected, but increased in the south (Bishop and Cook 1980, Jones 1982). ... On the other hand, melanics in south Wales were better camouflaged than typicals, yet comprised only about 20% of the population (Steward 1977a,b). Compiling data from 165 separate sites in Britain, Steward found a correlation between melanism and the concentration of sulfur dioxide (an airborne pollutant) north of latitude 52*N (Figure 2), but concluded that "in the south of Britain non-industrial factors may be of greater importance" than selective predation. Steward cautioned that "it may not be possible to generalize from the results for one area, to explain geographic variation over the rest of Britain" (Steward 1977a, pp. 239, 242).(d) The frequency of typicals on the Wirral Peninsula increased dramatically before the return of lichens to tree trunks (Clarke et al. 1985, Grant et al. 1998). ...(e) The decline of melanism in The Netherlands has been accompanied by an increase not only in typicals, but also in an intermediate form almost as dark as melanics. (Brakefield 1990). ... After the passage of anti-pollution legislation, the proportion of melanics decreased north of London (as expected), but inexplicably increased to the south (Bishop and Cook 1980; Jones 1982). In The Netherlands, the decline of melanism took another twist. As air pollution declined, not only did the frequency of typicals increase, but also the frequency of an intermediate form which was almost as dark as melanics, suggesting a more complex change than was seen in Britain (Brakefield 1990). |
||||
|
|
|
|||
|
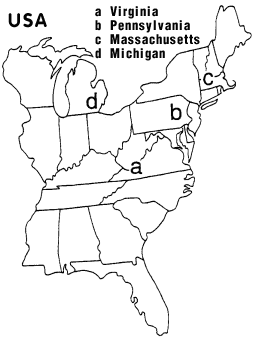
Problems with Pepper Moth Theory The U. S. A. |
U. S. A. |
|||
|
Map Key: |
Map and comments from: Second Thoughts about Peppered Moths, Jonathan Wells, Ph.D., 1999 |
|||
|
(b) From 1971 to 1978, melanics remained at about 52% in a low-pollution area in central eastern Pennsylvania (Manley 1981). ... In the United States, the first melanic peppered moth was reported in Philadelphia in 1906, and melanism increased until by 1959 it was over 90% in some areas (Owen 1962). ... Yet the geographical distribution did not fit the classical story any better than it did in the U.K. In a low-pollution area in central eastern Pennsylvania, melanics remained at about 52% from 1971 to 1978 (Manley 1981).(c) Melanics occurred at low frequencies in western and central Massachusetts even though trees were neither devoid of lichens nor blackened by soot (Sargent 1974, Treat 1979).(d) Between the 1960's and 1990's, melanics in southeastern Michigan increased and decreased in parallel with those in England, but without any perceptible changes in lichen cover (Grant et al. 1996). ... In southeastern Michigan, the frequency of melanics was over 90% in the early 1960's, then dropped to less than 20% by 1995 (Grant et al. 1995), thus paralleling the decline of melanism in the U.K. Yet the decline in Michigan "occurred in the absence of perceptible changes in local lichen floras" (Grant et al. 1996, p. 351). |
||||
|
|
|
|||
|
Pepper Moth theory falsified: No correlation between melanism and lichen cover but a correlation exists with pollution levels.
|
Recently, Grant and his colleagues reported a good correlation between sulfur dioxide levels and melanic frequencies in southwestern Virginia, central eastern Pennsylvania, and southeastern Michigan. In fact, the decline of melanism in both the U.S. and the U.K. appears to be "correlated primarily with reductions in atmospheric sulfur dioxide" (Grant et al. 1998, p. 465). The lack of correlation between melanic frequencies and lichen cover, however, is a serious problem for the theory that industrial melanism is due to cryptic coloration and selective predation. This aspect of the story warrants a closer look. Is there a correlation between melanism and lichen cover? In the 1970's, Kettlewell noted that melanism began declining on the Wirral Peninsula before lichens returned to the trees (Kettlewell 1973). Lees and his colleagues surveyed melanism in peppered moths at 104 sites throughout Britain, and found a lack of correlation with lichen cover which they considered "surprising in view of the results of Kettlewell's selection experiments" (Lees et al. 1973). As noted above, American biologists during the same period reported that melanism was also unrelated to lichen cover in the U.S. (Sargent 1974, West 1977, Treat 1979). The discrepancy has persisted. In the early 1980's, Clarke and his colleagues found "a reasonable correlation" in the U.K. between the decline in melanism and decrease in sulfur dioxide pollution, but were surprised to note "that throughout this time the appearance of the trees in Wirral does not seem to have changed appreciably" (Clarke et al. 1985, p. 198). According to Grant and Howlett, if the rise of industrial melanism was originally due to the demise of lichens on trees, then "the prediction is that lichens should precede the recovery of the typical morph as the common form. That is, the hiding places should recover before the hidden. But, this is clearly not the case in at least two regions where the recovery of typicals has been especially well documented in the virtual absence of these lichens: on the Wirral..., and in East Anglia" (Grant and Howlett 1988, pp. 230-231). Ten years later, the situation on the Wirral Peninsula remains the same; Grant and his colleagues note that "even now lichen coverage on trees in that region is sparse, yet the typicals exceed 93%" (Grant et al. 1998, p. 466). The decline of melanism in the U.S. without perceptible changes in the lichen cover prompted Grant and his colleagues to conclude that "the role of lichens has been inappropriately emphasized in chronicles about the evolution of melanism in peppered moths" (Grant et al. 1996, p. 351). Sargent and his colleagues note that "the recent declining frequency of melanism in B. betularia in North America, where the hypothesis of a cryptic advantage of melanism never seemed applicable," is "perplexing" in view of the classical story (Sargent et al. 1998, pp. 316-317). So melanism declined in the U.K. before lichens returned to the trees. In the U.S., melanism occurred in the presence of lichens and declined without any perceptible change in them. Clearly, the rise and fall of industrial melanism did not depend on the presence or absence of lichens. Yet their presence or absence seemed highly significant in Kettlewell's experiments. Why the discrepancy? |
|||
|
|
|
|||
|
Footnotes from snippets of "Second Thoughts about Peppered Moths", Jonathan Wells, Ph.D., 1999 |
||||
|
|
|
|||
Book Critique:
What does Majerus think about the Textbook theory:
Majerus rejects the textbook story of the Pepper Moth, but still holds to the conclusions of textbook story even though most of its component parts are wrong and others quite uncertain. In throwing his support to the "selective bird predation" theory of the Pepper Moth, it is clear that Majerus is making an educated guess. In fact, he concludes his discussion of the Pepper moth by quoting D. Futuyma on page 156 who says, "We don't know the whole story yet, But it should be possible to find out what it is." Then Majerus adds to Futuyma by saying in a less optimistic tone, "we may only have two decades". So although Majerus holds to the "selective bird predation" theory, he is not sure it is true and predicts that in short time, we will never know for sure. Majerus has offered us his opinion. Of course, it is our opinion that the evidence is so overwhelming against the Pepper Moth theory, that it has already been thoroughly falsified.
|
|
|
|
|
Melanics existed before industrialization: No new genetic information: Black, white moths and all shades in-between existed before and after. |
|
|
|
|
|
No single factor |
|
|
|
|
|
Multiple identical mutations "We doubt, however, that anything more is involved in these cases than the selection of already existing genes." (Fred Hoyle and Chandra Wickramasinghe, Evolution from Space, 1981, p. 5) |
Melanism induced by environmental agents (abiotic)
|
melanism induced by temperature and humidity |
Some entomologists noted that in experiments with butterflies, rearing at low temperature and high humidity produced melanic adults, and consequently they argued that melanic moths might be produced as a direct result of environmental factors. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p101) |
|
melanism induced by metallic salts |
After the First World War, JW. Heslop Harrison (1920) reviewed previous work on melanism. He specifically rejected Tutt's (1896) explanation based on differential bird predation, favouring instead the view that melanic forms arise as a consequence of metallic salts in pollution fallout on to leaves, which are eaten by larvae. ... In brief, these appeared to show that such treatment could lead to the production of melanic forms after a few generations. ... Porritt (1926) criticises Harrison's experiments at some length, commenting: ... Porritt's main criticism was that the species in which Harrison obtained a response frequently did not show melanism in the wild, even in industrial regions, where Porritt maintained that their larvae must be feeding on leaves polluted with the salts Harrison used. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p103) |
|
Melanism induced by various pollutant gasses |
Hasebroek (1925, 1929) conducted experiments of a similar type, but using pollutant gases, such as methane, ammonia, pyredin, chloroform, and sulphuretted hydrogen vapour, that might be absorbed through the tracheal system of pupae. His results from work on a dozen species of moths and butterflies showed that some individuals did show a darkening of the wings. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p103) |
The Kettlewell experiments
|
At this point, it is only pertinent to describe the main experiments that established beyond reasonable doubt that differential predation of the two forms by birds was the principal cause of the rapid spread of the carbonaria form. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p104) |
|
|
Kettleworth reported key prediction proven true.
(black moths dominate polluted areas)
|
On the basis of this connection, one should be able to predict, at least roughly, the frequencies of melanic and non-melanic forms in different geographic regions, on the basis of pollution levels and their influence on habitats . The carbonaria form should attain high frequencies in heavily polluted areas, while f. typica should still predominate in unpolluted regions. Areas with intermediate pollution levels should have both forms present, at intermediate frequencies. ... The various surveys conducted during the 1960s and 1970s allowed frequency maps to be drawn and considered alongside maps showing pollutant concentrations (Figs 5.4 and 5.5). Such comparisons suggested a very strong correlation between melanic frequency and pollution levels. High melanic frequencies occurred primarily close to, or to the east of, industrial centres. Rural regions in the west of England and Wales, and the north of Scotland were characterised by an absence, or very low frequency of carbonaria. The high frequencies in apparently rural parts of eastern England were explained by Kettlewell as the result of the prevailing south-westerly winds blowing pollutants across from the industrial Midlands and London. This explanation seems convincing given the scarcity of lichens on tree trunks in the eastern counties. Indeed, in 1954, when Kettlewell was seeking an unpolluted woodland as a site for predation experiments, he states that he could find no lichened woods in the eastern half of England south of Yorkshire. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p111,112) |
|
Pepper moths do not rest on tree trunks, but hidden high up underneath lateral branches.
|
|
|
Prediction based upon faith. It may be that birds rarely find wild pepper moths if they rest underneath lateral branches. Such a find would completely falsify the pepper moth story as an example of evolution. |
Although observations of peppered moths being taken from natural resting positions are still lacking and are urgently needed, it is highly probable that predation levels are significant. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p125) |
|
Birds eat significantly less moths (if any at all) when in their native resting position high in trees underneath lateral branches. Howlett and Majerus' recorded a 25% reduction in 1987 by placing moths on the trunk below branch joints, while Jones a 50% reduction in 1993 by placing the moths "under lateral branches", clearly proves our point. |
|
The data sets from Howlett and Majerus' (1987): Reduction of about 25%
|
Note: divide percentages by two to get actual numbers of moths (From Howlett and Majerus 1987.) |
Typica (light) |
carbonaria (dark) |
|
|
Eaten |
Eaten |
||
|
Stoke-on-Trent (dark) |
Exposed Trunk |
58% |
40% |
|
Trunk/branch joint |
50% (13% less than exposed) |
28% (30% less than exposed) |
|
|
New Forest (light) |
Exposed trunk |
32% |
62% |
|
Trunk/branch joint |
26% (25% less than exposed) |
40% (35% less than exposed) |
|

|
Steward concludes birds are not the primary factor of melanism. Notice that Majerus claims that Steward's conclusion is unsound. But the mortality rate of moths below lateral branches will be significantly less than for moths on the trunk where Steward placed them. Majerus' correct criticism of Steward reduces the importance of birds, not increase them. If Steward had placed the moths under lateral branches, he may have rejected bird predation as a factor all together!
Click on photo to see high resolution The Diurnea Fagella (melanic form) unlike the Pepper Moth, is a trunk rester that falsifies the Pepper moth theory. |
Steward (1977a) investigated the relationship between predation and morph frequency further by an indirect method. He assessed the 'relative crypsis' of the forms at 52 sites in southern England and compared the values obtained with the frequencies of the forms. Although he found that there was a correlation, it was weak, only accounting for 18 per cent of the variation in the frequencies in carbonaria between different sites, and 3 per cent of the variation for insillaria. ... He concluded that for southern Britain, selective predation is only of secondary importance with regard to melanic frequency of the peppered moth, some other factor being the main determinant of melanic frequency. For two reasons Steward's (1977a) conclusion must be deemed unsound. First, unfortunately, Steward assessed relative crypsis of moths by comparing them with tree trunks. While this is reasonable for the green brindled crescent and Diurnea Fagella which both naturally rest on tree trunks, it is not reasonable for the peppered moth, which rests elsewhere. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p132) |
Birds see in Ultra Violet, humans and moths do not.
|
This applies only to unpolluted forests with lichen: Humans and birds see pepper moths similarly in both normal and UV light. But humans and birds see the resting places of moths differently in UV.
The theorized (textbook story) resting place of Typica is on foliose lichens.
Black pepper moths on foliose lichens [light] are easily seen in human vision, but in pure UV black moths appear camouflaged on foliose lichens.
Typica (light moths) are best camouflaged on foliose lichens in human vision, but in pure UV the opposite it true. Conversely black pepper moths are camouflaged on foliose in pure UV but the reverse is true in human vision.
|
the visual acuity of birds :As I have already pointed out (p. 126), birds do not see as we do, having greater discriminatory prowess, and being highly sensitive to UV wavelengths, to which we are virtually blind. The exact way in which birds use their UV sensitive cones is not known. However, it seems more likely that they effectively have tetrachromatic vision, in which all four types of cone are stimulated simultaneously; thereby producing a mean wavelength from any point being viewed, rather than having the ability of using the UV sensitive cones independently from their other cones. If this is the case, it might be argued that as Kettlewell (1955a) and Steward (1977a) were mainly interested in the relative crypsis of the forms of the peppered moth in particular places, what birds actually see compared with humans does not actually matter as long as the differences are the same for both forms of peppered moth and for the backgrounds upon which they rest. This argument only holds if the UV reflectance characteristics of typica and carbonaria, of lichens of various species and of the bark of appropriate trees are the same. This is not the case. Jim Stalker, has assessed the UV reflectance characteristics of all the common forms of the peppered moth, together with six lichen taxa taken from seven tree species. He found that while the black scales and hairs of all forms of the peppered moth absorb UV wavelengths, the white scales reflect UV strongly. Stalker's results in respect of lichens are of particular interest. A number of authors have stressed the correlation between the frequency of typica and foliose or vegetative lichens. Stalker thus considered foliose and crustose lichens separately. All parts of the foliose lichens analysed, including Hypogymnia and Evernia spp., absorbed UV. Conversely, some parts of the crustose lichens examined, e.g. Lelicanora spp., reflected UV, other parts absorbing it giving an overall speckled appearance in pure UV. This meant that, although carbonaria was very obviously more conspicuous [more visible] than typica in the human visible spectrum when the two were set against bark covered in foliose lichens [light to humans, black in UV], the reverse was true in the UV spectrum. On the other hand, when set against bark covered with Lelicallora conizaeoides [speckled in UV], typica was less visible than carbonaria both in human visible and UV wavelengths (Stalker ef al. in preparation). Stalkcr's results led to two deductions. First, none of the assessments of the relative crypsis of moths as determined by humans should be applied to birds. Second, it seems likely that the patterning of the typical form of the peppered moth has evolved to be cryptic against crustosc lichens, not foliose lichens. If this is so, it seems probable that the ancestral daytime resting site for the peppered moth is under horizontal branches, which in unpolluted regions are usually encrusted with crustose lichens. The moths are unlikely to rest on the upper surfaces of the branches, as these tend to support a rich foliose lichen flora (Liebert and Brakelield 1987). (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p136) |
Do dark moths prefer dark resting backgrounds and light moths prefer light?
The only way the pepper moth theory can work is if moths randomly select light and dark resting places regardless of their wing colour. If moths chose resting sites that matched their wing colour, then they would always be camouflaged and "selective bird predation" would not be a factor. The fact seems to be proven that all moths generally prefer darker backgrounds. (should this really surprise us?) But even this falsifies the pepper moth theory because light pepper moths would chose dark coloured resting spots in unpolluted forests, making them easy prey for birds. In other words, light Pepper moths would not chose resting sites on light tree trunks, as commonly pictured in school textbooks.
Majerus' own data:
|
Table 7.9 Deductions from field records, observed releases, and laboratory experiments on natural resting positions and rest site preferences of some British Lepidoptera (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p188) |
|||
|
Species |
Probable common resting positions |
Species specific rest site selection |
Morph specific rest site selection |
|
Mottled beauty |
On tree trunks and under branches |
prefers black |
Banded rests more often across black/white border (compared with other forms). E nigra and f. nigri cata versus non-melanic: no difference |
|
Willow beauty |
On tree trunks and under branches |
prefers black |
melanic stronger preference for black than non melanics |
|
The results of these detailed investigations demonstrate that the morph frequencies of many species of Lepidoptera depend crucially upon habitat type. In 14 of the 19 species in which two or more morphs were taken in sufficient numbers to allow meaningful analysis, significant differences in the frequencies of morphs were recorded over a distance of 20m, when an abrupt change in habitat type occurred in this distance. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p189) |
|||
|
|
Resting site selection: Much has been written about the possibility that the different forms of the peppered moth behave in different ways when choosing resting sites. ... The idea that a mechanism will evolve which enables different forms of a moth to rest in different positions appropriate to their appearance is intuitively seductive. For example, melanic peppered moths may evolve to rest preferentially on relatively dark homogeneous backgrounds, while typica retains an ancestral behaviour of resting on paler and more speckly backgrounds. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p137) |
|
Kettlewell concluded that moths prefer to rest on surfaces that maximize their camouflage. But the most recent efforts have been unable to duplicate Kettlewell's results. Using light analysis, Howlett and Majerus predicted and confirmed that all pepper moths would prefer an artificial plain black surface. |
Kettlewell (1955b) first conducted experiments to determine whether the different forms of the peppered moth actively choose resting positions which maximize their crypsis. He placed peppered moths in barrels lined with equal surfaces of black and white card. His results showed that typica rested preferentially on the white surfaces, carbonaria preferring black (Table 6.4). Other workers have obtained similar results with the peppered moth (Boardman et al 1974; Kettlewell and Conn 1977) and other species (Steward 1985). However, others who have tried to replicate the experiments with the peppered moth have failed to obtain evidence of resting site selection (Mikkola 1984), or have obtained contradictory evidence (Howlett and Majerus 1987; C. Jones 1993). Howlett and Majerus (1987) analysed the light reflected from the wings of peppered moths. They showed that due to the partial translucence of the typical moth's wings, the wings are more similar to a plain black surface than to a plain white one when place against most natural surfaces. So, when presented with a choice of black or white surfaces, both forms should prefer the darker surface. Such tests were made and the expectation confirmed. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p138) |
|
Grant and Howlett found that some light and dark pepper moths were attracted to certain colour background, while others had no preference at all. |
More recently, Grant and Howlett (1988) demonstrated that some individual peppered moths do appear to have preferences for backgrounds of particular colours, but that these preferences are not correlated to the moths' phenotypes. In addition, some individuals had no preferences. The issue of whether and how the forms of peppered moth select appropriate resting positions is fraught with controversy and full of contradictory evidence. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p140) |
|
If Pepper moths possess the ability to match their wing colour to resting position then both colours would select camouflaged resting positions regardless of whether they are in polluted or unpolluted forests. This would completely falsify the pepper moth story because all moths would rest in camouflaged places regardless of the forest they were in. Remember, although the tree trunks were darkened by industrialization, these forests still have abundant resting locations to camouflage light pepper moths. |
First, the existence of a sophisticated resting site selection mechanism would reduce the importance of differential visual selection in the maintenance of the polymorphism (Cook personal communication). The habitats in which peppered moths live are heterogeneous. Dark homogeneous and pale heterogeneous backgrounds occur in all such habitats, whether in rural or industrial areas. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p140) |
|
This is one of the most proposterous sections of Majerus' book. We have Epicycle upon epicycle to explain the data. Faced with site selection results that are all over the map, he theorizes that the different experiments used moths that: 1. were dark colour but preferred light colour resting positions because they had not yet evolved the preference of dark. 2. were dark colour and had no preference being 1/2 evolved to dark preferences. 3. were dark colour and had fully evolved a dark colour resting preference. It would be far more credible to simply charge Kettlewell with falsifying his data than engage in such theories. We marvel that Majerus' would propose a theory that, if true, would explain why different site preference experiments produce contradictory results, but fully falsify the pepper moth story. This is because moths, in a state of transitional evolution, would land on sites that would cause them to be eliminated. So if the pepper moth has a site preference based on it wing colour, this preference must be fixed, stable AND CORRECT for dark and light coloured moths. Majerus does have the insight to recognize this for he says: "These results suggest that the selection of appropriate resting sites will only evolve if there is linkage between the colour pattern locus and the behavioural locus." (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p141) |
If this was the case, Kettlewell's results would be explained. The same theory could explain other apparently contradictory results. So, for example, the results of Howlett and Majerus (1987) in which both typica and carbonaria moths showed a preference for black surfaces would be explained if the moths were from a population in which the dark allele had been fixed. It is perhaps relevant to note that the moths used by Howlett and Majerus came from the Cambridge area, at a time when a frequency of carbonaria was about 40 per cent, and was declining from a level of over 90 per cent in the late 1950s. In trying to unravel the causes of the great variation in the results of back-ground choice experiments in the peppered moth ... The problem is therefore unresolved. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p143) |
|
If light moths had developed the mechanism to rest on matching backgrounds, but then evolved black wings, they would still land on light backgrounds resulting in their detection by birds. This is a major problem and Majerus knows it. |
In trying to unravel the causes of the great variation in the results of back-ground choice experiments in the peppered moth ... The problem is therefore unresolved. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p143) |
|
Majerus theorizes that all pepper moths chose dark resting areas. We suggest that this is probably the truth and agree with Majerus's theory. But this would falsify the pepper moth story because the camouflage value of light and dark moths is quite similar in dark hiding places. Hence the relative camouflage efficiency for both colours of moth is high, resulting in an overall reduction of predation by birds AND a reduced selective choice on those rarer occasions that birds do find a moth to eat. Stated differently, the relative contrast between black and white reduces proportionately with the light shining on them. Lower light, lower contrast between white and dark objects. To illustrate, look at a stack of books on a bookshelf with the lights on, then with the lights off. In lower light conditions the difference between black and white reduce. Same for the pepper moth when both light and dark coloured moths select dark resting places between or under leaves and twigs or under bark etc. |
Thus, if the peppered moth is to express a resting site preference in artificial cylinder experiments, it might be expected to select a dark surface, reflecting its natural resting position on trees. This preference would be independent of morph as the crypsis of all forms of the moth would be maximised by resting in shaded areas. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p146) |
Do light mate with light and dark with dark?
|
If light mate with light and dark with dark, indeed it would explain why the population colours become fixed. However humans generally mate according to skin colour in multicultural environments and such facts do nothing for evolutionary theory. But if moths do not mate with like colours, it falsifies the pepper moth story because the gene pool would always posses a mix of light and dark coloured genes. |
Effect Of Mating On Selective Predation: Work to test the effect of mating with a similar or different morph on levels of selective predation is urgently needed. Not only will such work help to increase the accuracy of estimates of morph fitness, but a finding that morphs do tend to mate assortatively (like tending to mate with like) would be of great significance in respect of general evolutionary theory. If it were shown that a preference to mate with a similar rather than with a different phenotype had evolved because the mating between different forms was at a selective disadvantage, this would perhaps represent the first stage in the process of speciation through disruptive selection. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p146) |
Melanism in one sex of a moth species
According to the pepper moth theory, the Ghost Moth should be wiped out by eliminating one sex by selective bird predation and the Mulsin Moth, which is not eaten by birds, cannot be explained.
Darwin commented on the Ghost moth that has a bright white male and a dark coloured female.
The Buff ermine is not eaten by birds but has a melanic form:
|
Buff Ermine Typical (spilosoma luteum)Here we have a moth, the Buff Ermine, that is not eaten by birds because of it toxicity. Obviously selective predation by birds was not a factor in this case of melanism. Although Majerus admits the moth is unstudied, he ventures to guess that the moth evolved into the melanic form in order to not be detected by predators. But if the moth is not eaten by birds, what predators are we talking about? He sort of makes it appear that the moth had to sit down and weigh the benefits of such a change then decide to trade off its aposematic coloration for camouflage. However such a change is not even needed because the typical moth survives just fine! Evolution theory needs many epicycles to make it work! |
While melanism in the ruby tiger occurs over a wide area, that in the buff ermine, Spilosoma luteum Hufnagel, is a much more local phenomenon. The buff ermine is normally yellow, lightly spotted on both wings and abdomen, with black. It is relatively unpalatable, but is not as toxic as some other members of the genus (Rothschild 1963). In a few coastal sand-dune locations, on the east of England, the continental Channel coast and some of the islands off Germany, a melanic form, f. zatima, occurs as a polymorphism. This form has most of the intervenic regions of all the wings dark brown, with just the veins and small areas of the forewing retaining the normal yellow coloration. The zatima form is controlled by a single allele, showing incomplete dominance, the heterozygote being less extreme than the homozygote. Although little work has been conducted on the adaptive significance of this form, it is probable that in the locations where it occurs, its colours are cryptic. On the Yorkshire and Lincolnshire dunes, it appears to rest on the dead trunks and branches of elder, against which it is very well camouflaged. Such cases, in which aposematic coloration has been forfeited for the advantages of not being detected by predators, are rare. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p210) |
Hundreds of melanic moths (more epicycles) that selective bird predation of the Pepper moth theory cannot explain.
|
Majerus discusses a number of moths that he admits had melanic forms that predated industrialization. An important fact to remember! An a huge epicycle for Pepper Moth theory advocates!
|
Partial industrial melanic polymorphism: Perhaps more species of British moth fall within this category of melanism than any other. Most species that had melanic forms prior to the industrial revolution fall into this class, if their distribution includes major industrial regions, for in the majority of such cases, the frequency of melanics increased in the nineteenth and first half of the twentieth centuries. The case of the willow beauty, mentioned above, demonstrates this principle. Originally a non-industrial melanic, f. perfuniaria, rapidly increased in frequency in and around London, in the second half of the nineteenth century, appearing in other industrial heartlands such as Sheffield in 1902, Newcastle upon, lyne in 1914 and Burnley in 1916 (C. Jones 1993). Other similar cases include the green brindled crescent (Plate 4e and f), the clouded-bordered brindle, Apainea crellata (Hufnagel) (Plate 4g and h), and the satin beauty, Dedeptenia ribeata (Clerck), as well as most of those species showing morph specific habitat preferences discussed previously (p. 184). (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p202) |
|
What we have here proves how silly and unscientific the whole pepper moth theory is. Many moths have light and dark variations, always have, always will. There were dark and light moths before and after the "industrial pollution" blackened the trees. Most of the moths that have melanics are not related to "industrial pollution". Even Majerus admits he cannot prove that selective bird predation is the key element in the pepper moth theory. So of all the 13 moths Majerus lists (in fact there are hundreds that have dark and light variations), none of them experienced the kind of change noted in the pepper moth. Two of the 13 moths had a slight increase of darker moths in polluted areas. But 11 moths species did not! How unscientific to select only that small data that you want to highlight. Of these two species of moths that had a few more darker moths in polluted industrial areas, Majerus has the nerve to suggest that it was "probably as a result of increased crypsis" [camouflage] . So Majerus highlights the "common marbled carpet and clouded drab" as proof of the Pepper moth theory in action but ignores the other 11 moths where moths experienced decreased crypsis or camouflage. |
Several of the commonest species of British moth have a wide variety of different forms, including melanics. They include: the common marbled carpet; grey pine carpet; July highflyer; mottled umber (Plate 5b and Fig. 7.16); large yellow underwing; clouded drab, Orthosia incerta (Hufnagel); beaded chestnut, Agrochola lychnidis (Denis and Schiffermijiler); dunbar, Cosmia trapezina (Linnaeus); common rustic Mespainea secalis (Linnaeus), and lesser common rustic, Mespamea secalella (Remm). In all of these species, melanic forms occur in non-industrial regions, although in some, such as the common marbled carpet and clouded drab, melanics occur at higher frequency in industrial regions, probably as a result of increased crypsis. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p197) |
|
Majerus confesses the truth that the pepper moth almost stands alone as an example of "pollution that darkened tree trunks which caused birds to eat light moths on the dark trunks because of reduced camouflage eating" shift in colour from light to dark. Notice the only examples Majerus can give already had melanics before industrialization and the shift to melanic happen in the 1970's, a full 100 years to late. If the Pepper moth story is true, then why would melanics of these species develop 20 years after the forests began to be less polluted? Can't be because of a lack of genetic information needed to cause the change from light to dark because Majerus admits the genetic information pre-dated industrial age.
|
The classic example of full industrial melanic polymorphism is, of course, the peppered moth. The number of other examples of British moths that fall into this category is rather small . These include a number of species in which melanic forms had been described from single individuals caught in non-industrial regions, before the establishment of true genetic polymorphisms (sensu Ford 1940a). A case in point is that of the great prominent, Peridea anceps Goeze, in which melanic form, f. fusca, has been recorded from Westmoreland (now Cumbria). This form appears to have been inherited as a unifactorial recessive. More recently, a phenotypically similar form, inherited as a unifactorial dominant, has established as an industrial melanic in southern England, particularly around London, where the frequency of the melanic form reached over 40 per cent, in some localities, in the 1970s. Of course, it is not always easy to be absolutely certain that species with melanic polymorphisms, in areas affected by industrialisation, did not have such forms maintained at low frequencies prior to industrialisation. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p197) |
|
Remember, Majerus is painting with a very narrow brush when he says that some species are observed to increase their melanic populations during the industrial age. In fact, there were 10 times the moths that did not fit the theory. Majerus makes the incredible statement that although the genetic information needed to produce dark and light moths existed in other parts of Britain, they arose by mutation rather than migration, in new areas. England is a tiny land mass. It is obvious that the melanic moths started showing up in new area of the England because of migration, but to an evolutionist, everything must caused by multiple identical mutations! We reject this mutation theory in favour of the simplicity of migration a few hundred miles. |
There is no doubt that in some species, melanic forms that existed as polymorphisms before the widespread industrialisation , have increased in industrial areas subsequently. However, in other cases, new melanic forms have arisen by mutation and then spread in industrial regions of Britain, despite the existence of melanic forms in other parts of Britain. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p198) |
|
Majerus believes it was "serendipitous" (dumb luck) that melanism didn't develop earlier. Websters defines "serendipitous" as originating from a fairy tail where a princess has an "aptitude for making fortunate discoveries accidentally". But after 1950, the forests became less polluted and lighter, favouring the non-melanics. This is not dumb luck but bad luck for they developed at the wrong time. But most important, we note that selective bird predation cannot account for why these moths increased their melanic populations after 1950. |
In a few species, industrial melanism has developed very recently, within the last half of this century . This is true of both the sprawler, Brachionycha sphinx (Hufnagel), and the early grey, Xylocampa areola (Esper), in which melanic forms have increased considerably over the last three decades. The reasons why industrial melanism did not develop in these species earlier is probably serendipitous: the relevant mutation simply did not arise in the right place at the right time. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p198) |
|
Majerus' comments regarding the Pepper moth and the Oak Beauty are so preposterous we simply cannot contain ourselves . More epicycles! He argues that the Oak Beauty had not developed the melanic mutation on English soil, and this is the reason why the population did not increase in melanics. However, Majerus tells us that the mutation already existed in the Netherlands! Of course if you look carefully at Majerus' comment, the "mutation" had happened on English soil, "but has never occurred at appreciable frequencies in Britain". So there were melanics on British soil and the Netherlands before, during and after industrialization, but in smaller numbers. Majerus knows that the oak beauty is proof that the pepper moth theory is falsified by the Oak Beauty's failure to increase in the proportion of melanics. Remember, the Oak Beauty, is the pepper moth's closest relative!
|
These recent cases of industrial melanism emphasise an important feature of evolution by natural selection. Natural selection cannot cause change unless it has phenotypic variation to work upon. If a melanic form does not arise by mutation (or recombination), in an area where it would be at an advantage, selection has nothing to act upon. The case of the peppered moth's closest British relative, the oak beauty, Biston strataria (Hufnagel) (Fig. 7.17), is instructive in this regard. This species has a melanic form, f. melanaria, which is a recent and common industrial melanic in the Netherlands, but has never occurred at appreciable frequencies in Britain. To date, L melanaria has only been found here as a rare mutation, from Kent, Surrey, Hampshire, and Cambridge. The ecology and behaviour of the oak beauty is similar to that of the peppered moth in many ways. However, the melanic mutation in the oak beauty seems never to have arisen in this country in favourable circumstances, with the consequence that it has not successfully established itself here. In the Netherlands, f. inelanaria seems to be analogous to f. carbonaria. Other forms of the oak beauty also occur in the Netherlands. Two of these, f. nigricalls and f. robiniaria, both have wings suffused with dark brown, and perhaps parallel the darker forms of the insularia complex in the peppered moth. Both these forms occur in Britain, but never at high frequency. The relatively recent spread of f. melanaria in the Netherlands, in conditions that appear to mirror those experienced in Britain, and the lack of industrial poly-morphism in this species in Britain, emphasises the importance of mutational events occurring in the right place and time. Given the reversal of the fortunes of melanic peppered moths since the 1960s, it is unlikely that the melanaria form of the oak beauty will establish itself in Britain in the foreseeable future. In at least one moth, industrial melanism in Britain is the result of migration from the continent. The melanic form, f. fusca Cockayne, of the figure of eighty (Plate 4a and b) appeared in southern and eastern England in the mid-1940s, and has increased rapidly since, so that over much of its range in England, f. fusca is currently more common than the non-melanic form. Although f. fitsca is found in Wales and the Channel Islands, it has yet to be recorded in Scotland or Ireland. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p198-199) |
Industrial melanism in the Kola Peninsula?
Mikkola did not document that the melanic f. inferna, increased in the Kola Peninsula, only that he discovered a moth he "interprets" or suspects fits the Pepper moth theory. Even if the melanic form did increase in numbers, studies have proven that moths exposed to pollution will darken in colour. Hey, the area is so polluted the trees are dying off! Eero Tikkanen and Irja Niemelä edited a major report called, "Kola Peninsula pollutants and forest ecosystems in Lapland".
|
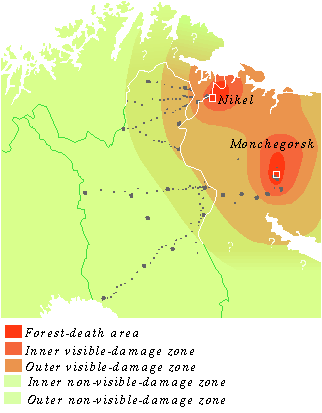
This report says in its conclusion: "The inner visible-damage zone extends ca. 30 km south and ca. 40 km north of Monchegorsk." Click on photo for high resolution "Figure 83. The forest-death areas in the vicinity of Monchegorsk and Nikel and the visible-damage and non-visible-damage zones in the Kola Peninsula and Finnish Lapland." |
Click on photo for high resolution "The conifers within this zone show marked defoliation and there are no epiphytic lichens to be seen. Severely defoliated pines ca. 30 km south of Monchegorsk. The crowns of the trees are more defoliated on the side facing the Severonikel metal smelter." ("Kola Peninsula pollutants and forest ecosystems in Lapland", edited by Eero Tikkanen and Irja Niemelä.) |
|
What Mikkola discovered was a moth that had a melanic form in the geographic area of the Kola Peninsula. There are other melanic moth species in the area. Mikkola did not say that he had documented an increase in the frequency of the melanic form of this moth, just that he discovered it and assumed it was another "Pepper moth" case. Yet selective bird predation is not even a factor! The area is so polluted that the trees are actually dying off! To single out one moth species from this area (f. inferna, of the noctuid Xestia gelida) and suggest it is a repeat of the Pepper Moth story, is simply untrue. Even if the melanic of this moth did increase, studies have proven that moths exposed to pollution will darken in colour. (see elsewhere in this file) But the death nail for Mikkola, is that the affected area is so small that migration of typical moths into the polluted area would offset the effect of the bird predation. (Should it ever be shown that selective bird predation is a factor here.) |
Although there appears to be a general decline in the frequency of truly industrial melanics; in Britain at present, this is not true in all other parts of the world. In Finland, for example, Mikkola (1989) discovered a new melanic form, f. inferna, of the noctuid Xestia gelida (Sparre Schneider), which he interprets as the first case of industrial melanism in the subarctic, and which lie associated with the intense pollution emanating from the Kola Peninsula. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p201) |
Majerus' "search image" theory (epicycle) flip flop.
Majerus' invokes the "search image" theory to explain why melanics (when their numbers are small) are not eaten by birds in unpolluted forests. Then he flip flops with was we call the "oddball target" theory that he uses to explain why melanics (when their numbers are small) are eaten by birds. Shouldn't the net effect be zero? Here we have two epicycles working against each other!
|
"The search image theory" that Majerus dreams up like a "Ptolemy epicycle" to explain that which doesn't fit the Pepper Moth theory. Here is the search image theory in simple words: Lets say a moth has 95% light and 5% dark in a forest with light coloured tree trunks. The theory states that under these circumstances the dark moth on a light tree is less likely to be eaten than a fully camouflaged moth merely because the birds are not familiar with the dark moth. Notice that this theory suggests that birds will eat one species of dark moth it is familiar with but not eat an unfamiliar species of dark moth. Problem is that this theory is as ridiculous as it is wrong. Birds are already looking for light and dark moths in any forest because there are hundreds of species of moths of every colour. So with 25 species of dark moths already being eaten by birds, one more won't change things. The "search image" theory suggests that although birds are already hunting for and eating 25 melanic moths, that if a new one shows up, they will continue to eat the 25 they are familiar with but reject the 26th. Very unscientific and most unbelievable. This theory is involked whenever the data doesn't fit the standard "pepper moth theory" to explain away anomalies in data which only epicycles can harmonize.
|
|
|
The "oddball target" theory: Here Majerus completely flip flops on the "search image" theory. Now he says that birds will try the rare oddball moth, whereas the "search image" theory suggested that birds would not try such a moth because they are not looking for it. More epicycles. |
Furthermore, given the general theory of the selective advantage of aposernatic coloration, it is likely that completely black forms of these species will be maladaptive in the wild, because of their rarity. Birds that have learnt to avoid the common typical forms of these species are unlikely to avoid forms that are so different, without at least trying them. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p209) |
Very weak support for the Pepper Moth theory in these three moth species.
Why is it that of all the moths that have melanic forms, the Pepper Moth is the only one whose melanic populations shifted from the minority before industrialization to a majority?
|
When Majerus says the frequency of melanics within the pale brindled beauty, the mottled beauty, and the green brindled crescent, increases in industrial regions, he is trying to echo the pepper moth theory with these 3 moths. But according to Majerus' own data, there are more typical mottled beauty's than melanics. So when he says an increase, he means from say 33% to 35%, but still in the minority. Notice that he even goes on to Green Brindled Crescent as a moth that is unstudied.
|
Rather few of the species that fall into this class of melanism have been investigated in any depth. Exceptions are the pale brindled beauty (Lees 1971, 1974, 1975) (Plate 4a and b), the mottled beauty (Kettlewell 1973; C. Jones et al. 1993) and the green brindled crescent (Steward 1977d). All three of these are trunk resters, and the increase in melanics in industrial regions is attributed to increased crypsis. ... Many species have received virtually no attention, and their status is certainly open to conjecture. I will give just a handful of examples, which would repay attention. Aniong the Noctuids, the brindled green, Dryobotodes erendta (Fabricius), has a recurring melanic form around London, and Skinner (1984) mentions that the darker form occurs frequently in the Midlands.(Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p203) |
Open and closed canopy experiments (20 meters apart)
Majerus accidentally discovers he needs to add more epicycles
|
Click on photo for high resolution |
Majerus writes: "I was extremely surprised to find that the proportions of melanic forms of two species, the mottled beauty and the tawny-barred angle, Semiothisa liturata (Clerck) (Plate 6d), were quite different between the two traps, although the traps had been set up less than 20m apart (Table 7.8)." (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p184) |
Majerus' Data:
|
Table 7.8 Details of the numbers of different forms of mottled beauty and tawny-barred angle, taken from Heath moth traps in different habitats, on 12 July 1984, at Ynys-Hir, Dyfed. Trap I was set up in mixed deciduous woodland, trap 2 in a Douglas fir plantation. The traps were set 20 yards apart. (From data in Kearns and Majerus 1987.) (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p184) |
|||
|
Mottled beauty |
Trap 1 |
Trap 2 |
Totals |
|
typica |
41 |
45 |
86 |
|
conversaria |
0 |
11 |
11 |
|
nigricatalnigra |
0 |
2 |
2 |
|
Totals |
41 |
58 |
99 |
|
Tawny-barred angle |
|
|
|
|
typica |
8 (87%) |
16 (20%) |
24 |
|
nigrofulvata |
1 (13%) |
20 (80%) |
21 |
|
Totals |
9 |
36 |
45 |
Why this is so devastating to the Pepper Moth theory:
Pepper Moth Theory Falsified:
Birds eat equal number of dark and light moths|
Majerus confesses that for the vast majority of moths that have both melanics and typicals, birds were indifferent on which one's they ate in both open and closed canopies. If you look at Majerus's data in the "appendix table A2", that summarizes the data into percentages, there appears to be slightly more dark moths in the darker forest (shaded by canopy not pollution). However, the overall ratio of dark and light moths is insignificant and nothing approaching what is needed to explain the Pepper moth theory. |
Observations of moths during these experiments [open vs. closed canopies] indicated that the main predators were birds, but ground beetles and ants also took some of the moths. In the majority of species there was no evidence of any significant difference in the level of predation on different morphs of a species in a particular habitat. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p187) |
|
Although Majerus highlights four moth species that fit his theory, that bird predation is a force of natural selection, there were more that did not. In truth he confesses, "However, in most instances the differences were not significant". If you look at the data yourself, some moths supported his theory, some moths contradicted his theory.
|
Broadly, three general conclusions were drawn from the predation experiments. First, in virtually every case, the level of predation was higher in the more open of two contrasting habitats. Second, in a small number of tests, involving the mottled beauty in Dyfed and Surrey; willow beauty in Hampshire; tawnybarred angle in Dyfed; dotted border in Hampshire and Suffolk and the small engrailed in Suffolk, the level of predation of melanic and banded morphs was lower than that of typical morphs in close canopy habitats, the reverse being the case in the more open habitats. However, in most instances the differences were not significant, and there was no strong consistency, either within or between species. Third, in the riband wave, the level of predation of the banded form was higher than that of the typical form in the closed canopy habitat, the reverse being the case for the open habitat. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p190) |
Selective bird predation experiments in closed and open canopies show that although birds eat more moths in open canopies, they do not prefer dark or light moths in any environment. This falsifies the whole "Pepper moth" selective predation Theory. Notice that the overall trend, although insignificant, is that birds almost always eat more non-melanic moths in both canopies. (3% more in closed and 1% more in open)
Majerus' own data:
|
|
Close canopy |
open canopy |
||
|
|
Melanic Eaten by birds % |
Non-melanic Eaten by birds % |
Melanic Eaten by birds % |
Non-melanic Eaten by birds % |
|
Mottled beauty (trunk rester) |
34 |
45 |
45 |
55 |
|
Willow beauty |
17 |
18 |
24 |
27 |
|
Tawny-barred angle |
15 |
19 |
24 |
22 |
|
Dotted border |
36 |
46 |
26 |
24 |
|
Common pug |
7 |
9 |
8 |
8 |
|
Engrailed |
22 |
20 |
31 |
24 |
|
Small engrailed |
16 |
19 |
19 |
16 |
|
Riband wave |
10 |
8 |
7 |
14 |
|
Average % |
20 |
23 (+3%) |
23 |
24 (+1%) |
|
Important note: |
Data above is simplified and combined from "Melanism: Evolution in Action", Michael E. N. Majerus, 1998, appendix. However the actual data presented here is not directly from Majerus, but based upon his appendix. Although we feel it is accurate, use caution therefore and buy his book yourself to check accuracy. |
|||
Statistics on actual numbers of melanic vs. typicals in open and closed canopies:
Notice that without exception, there are always more dark moths in closed (dark) canopies and more light moths in open (lighter) canopies. If the Majerus' original data is reliable, it poses a huge problem for the "pepper moth" theory because although we might conclude that the observed differences is the result of bird predation, specific experiments to prove this have failed. This means that according to Majerus' own data, "selective bird predation" is not the reason why there are more dark moths in closed (darker) canopies.
|
|
melanic (closed) % |
melanic (open) % |
Reduction of Melanic from closed to open canopy % |
|
Mottled beauty (trunk rester) |
36 |
15 |
21 |
|
Grey pine carpet |
83 |
61 |
22 |
|
Willow beauty |
26 |
10 |
16 |
|
Tawny-barred angle |
48 |
9 |
39 |
|
Pale brindled beauty (trunk rester) |
22 |
16 |
6 |
|
Dotted border |
11 |
2 |
9 |
|
Common pug |
89 |
78 |
11 |
|
Engrailed |
26 |
11 |
15 |
|
Small engrailed |
18 |
9 |
9 |
|
Riband wave |
29 |
16 |
13 |
|
Common marbled carpet |
66 |
53 |
13 |
|
Pine carpet |
43 |
30 |
13 |
|
Large yellow underwing |
52 |
20 |
32 |
|
Lunar underwing |
16 |
8 |
8 |
|
Average reduction |
|
|
16% |
|
Important note: |
Data above is simplified and combined from "Melanism: Evolution in Action", Michael E. N. Majerus, 1998, appendix. However the actual data presented here is not directly from Majerus, but based upon his appendix. Although we feel it is accurate, use caution therefore and buy his book yourself to check accuracy. |
||
Majerus summarizes his open/closed canopy study with 5 troubling conclusions:
|
This calls into question all of Kettlewell's experiments because he was not aware of this fact. for example if he had set his traps only 20 meters away, he would have collected contradictory data to the "Pepper moth" theory. |
The conclusions from this study may thus be summarised: (a) Correlations between morph frequencies and habitat type are common in the Lepidoptera and may produce large and very abrupt changes in morph frequency over a few metres across habitat boundaries. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p191) |
|
If you examine the data yourself, more species did not show correlations than those that did. Even worse, the moth species in (b) (c) and (d) are NOT THE SAME but different species. Such conclusions are unscientific. Look at the data. |
(b) In species showing morph habitat correlations, melanic and banded forms occur at higher frequencies in dark close canopy habitats, compared with lighter more open habitats. (c) In some species, differences in differential bird predation between habitats, occurring each generation, could be sufficient to explain the morph frequencies observed. (d) Some species exhibit morph specific habitat preferences of a magnitude sufficient to explain the morph frequencies observed. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p191) |
|
Majerus admits that the observed differences have nothing to do with bird predation . Although he says, "For some species", if you check the data yourself, it is in fact "For most species". Majerus has not misrepresented the data, but it is plainly obvious that what he is really saying by all this is that the data is "all over the map" and we cannot explain it. |
(e) For some species, no causative explanations of the abrupt changes in morph frequencies across habitat boundaries have yet been revealed. (Melanism: Evolution in Action, Michael E. N. Majerus, 1998, p192) |
|
|
|
|
Proof that textbooks in use this school year engage in classroom fraud. |
|
|
|
|
|
Evolutionary Biology, Douglas J. Futuyma, Third Edition, 1998 ISBN: 0-87893-189-9
In use April 2000 2nd - 4th year biology at McMaster University.
|
In order to infer the ADAPTIVE SIGNIFICANCE, or function, of the feature, the experiment should be done in the context of a postulated selective factor. The individuals whose performance or fitness is compared may differ either genetically or by experimental alteration. For example, H. B. D. Kettlewell (1955) suspected that the melanic (dark) genotype of the peppered moth (Biston betularia) had increased in frequency in Britain following the Industrial Revolution because, compared with the light form, it more closely resembled the color of tree trunks. Be-fore the Industrial Revolution, many trees were clothed with pale lichens, but air pollution killed many of these lichens, leaving bare, dark trunks. Kettlewell showed that when marked moths of both colors were released in a polluted woodland, a higher proportion of melanic than of typical moths were recaptured in subsequent days, whereas the reverse occurred when he released moths in a non-polluted wood. He confirmed that differential predation occurs by pinning moth specimens of both color forms on both bare and lichen-covered trees and recording the rates at which birds attacked them (Figure 12.23) [FIGURE 12.23 Wild-type (gray) and melanic (dark) peppered moths (Biston betularia) on (A) a lichen -covered tree trunk and (B) a trunk lacking lichens. The gray form is more cryptic in A, and the dark form is more cryptic in B. In one of Kettlewell's experiments, fresh specimens of both forms were pinned to both kinds of trunks. The more conspicuous form in each situation was removed more rapidly by birds. (Photographs by H. B. D. Kettlewell.)]. Typical moths suffered less predation than melanic ones on lichen-covered trees, whereas melanic moths had an advantage on dark lichen-free holes. (However, in both Biston and many other insects that display industrial melanism, predation is probably not the only selective factor that affects gene frequencies: Lees 1981). Andersson's (1982) alteration of the tail length of male widow birds, described earlier in this chapter, illustrates how artificially created variation may be used to demonstrate a feature's adaptive function, in this case its role in mating success. The interpretation of such experiments can be beset by many pitfalls (Endler 1986). (Evolutionary Biology, Douglas J. Futuyma, Third Edition, 1998 ISBN: 0-87893-189-9, p 358) Conversely, if the genetic constitution changes too fast to be accounted for by drift, selection is likely to be responsible. The most conspicuous such example is the famous case of "industrial melanism" in the peppered moth (Biston betularia). Museum collections dating since the mid-nineteenth century in England, less than a century after the onset of the Industrial Revolution, show a decline of the "typical" pale gray form and a rapid, rather steady increase of the black (melanic) form, from about 1 percent to more than 90 percent in some areas (see Figure 23 in Chapter 12). The rate of change is so great that it implies a very substantial 50 percent selective advantage for the melanic form (Haldane 1932). In this instance, we know further that the frequency of the melanic allele tends to be highest in heavily industrialized areas, where air pollution has killed off the pale lichens that invest tree trunks in nonpolluted areas. (Evolutionary Biology, Douglas J. Futuyma, Third Edition, 1998 ISBN: 0-87893-189-9, p 728) |
|
|
|
|
Biology, James M. Barrett, 1986, Prentice-Hall, p 754
|
"FIG. 27-12 Biston betularia, the peppered moth. The normal, white with speckled black, and the mutant black forms are shown on (a) lichen-covered and (b) soot-blackened tree trunks." ... "Variation: The Raw Material Of Evolutionary Change: As Darwin noted, there is great variation in structure, function, and behavior within each species. Although the evolutionary significance of many of these variations in heritable characteristics is unknown, some variations are adaptive in that they increase their possessor's chances of surviving long enough to reproduce under particular environmental conditions. For example, the appearance of a dark-colored variant of a normally light-pigmented insect in an industrial area blackened by soot may be of survival value to that population. In highly industrialized districts of England in which soft coal was used as fuel, the trees and other objects in the area gradually became darkened by soot. (Soot killed the lichens that normally lived on tree trunks, and consequently the dark color of the bark was exposed.) During the past 200 years, certain dark-colored mutants have appeared among a species of moth living in the industrial city of Manchester, England. Museum collections and records show that a dark variant of the peppered moth (Biston betularia) first appeared in Manchester about 1850 A.D. (Fig. 27-12). Collection records show that since that time, the dark mutant variety spread widely throughout the industrial belt of England. Unlike the mottled ancestral form, the dark mutant blends with lichen-free, dark tree trunks and is not as easily spotted by its predators (Fig. 27-12b). It is therefore more likely to survive long enough to repro-duce. Like all mutations, the dark mutation was a fortuitous occurrence, which under other circumstances might have been of no advantage to the moth; for that matter, against a light background, the mutation has been shown to be detrimental. Equal proportions of mottled and solid-colored moths were released in various environments and the survivors collected after a day or two. When the experiment was carried out in smoke-blackened regions of England, the percentage of mottled moths that survived was much lower than the percentage of dark-colored ones. Direct observations confirmed that birds were most successful in preying upon any moths that rested on sharply contrasting backgrounds, whether they were mottled moths on dark tree trunks or dark moths on lichen-covered, light-colored tree trunks. The mottled and the solid-colored moths are variant types of this species. The two variant types can interbreed if they coexist in the same region. However, each variant has better chances of surviving in a different type of habitat. These observations clearly show that a variant phenotype that is favored by a specific type of habitat becomes a pre-dominant or even an exclusive type in such an area. But in nature we often find a heterogeneous population coexisting within the same region. This heterogeneity may persist if the variation confers no survival advantage or disadvantage. In some instances, heterogeneity is maintained because natural selection favors the heterozygote. This is the reason for the persistence of the allele for sickle-cell anemia. (Biology, James M. Barrett, 1986, ISBN 0-13-076597-X, Prentice-Hall, p 754) |
|
|
|
|
Biology, Miller and Levine 2000 Edition
|
[Website Comment: Miller and Levine have made an effort to correct their textbook of the kinds of errors we are exposing on this website (www.bible.ca). However, Miller admits uncertainty of the most critical element of the pepper moth theory: Bird predation. Miller admits, "As Majerus explains, to be absolutely certain of exactly how natural selection produced the rise and fall of the carbonaria form, we need better experiments to show that birds (in a natural environment) really do respond to camoflage in the ways we have presumed ... Until these studies are done, the peppered moth story will be incomplete. Not wrong, but incomplete." (Ken Miller, Author of Biology by Miller/Levine, 2000 edition, from Browns university Website) We feel that Miller and other textbook authors would be well advised to restate it this way: "Until these studies are done, the peppered moth story will be incomplete. Not wrong, NOT RIGHT, but incomplete and unproven." Why teach as a central tenant of evolutionary theory that which we cannot be sure if it is right and in need of further study. Even if we had not falsified the Pepper Moth theory on this web page, any theory that is proven neither true or false is not science and does not belong in the science class room. Miller and Levine have an opportunity to be viewed by history as the first of the textbook authors who were honest and objective enough to remove the Pepper Moth story ALTOGETHER from the class room. Problem is, they know that there is little else to support evolution and they desperately cling to the story in hope that someone, somewhere will prove the story true!]Start quote below: "The Peppered Moth - An Update: The Peppered Moth is routinely used as an example of evolution. But is this well-known story wrong? For years the story of the peppered moth, Biston betularia, has provided one of the best-known examples of natural selection in action. The story of the moth was outlined on pages 297-298 of the Elephant Book, and highlights the field experiments of British ecologist H. B. D. Kettlewell. However, a recent book by Michael Majerus (Melanism -Evolution in Action) makes it clear that the peppered moth story will need to be rewritten. Joe Levine and I will post a revision of pages 297-298 here just as soon as we can, but in the meantime, here is an update of what the fuss is all about: ... However, Majerus also discovered that many of Kettlewell's experiments didn't really test the elements of the story as well as they should have. For example, in testing how likely light and dark moths were to be eaten, he placed moths on the sides of tree trunks, a place where they rarely perch in nature. He also records how well comoflaged the moths seemed to be by visual inspection. This might have seemed like a good idea at the time, but since his work it has become clear that birds see ultraviolet much better than we do, and therefore what seems well-camoflaged to the human eye may not be to a bird. In addition, neither Kettlewell nor those who checked his work were able to compensate for the degree to which migration of moths from surrounding areas might have affected the actual numbers of light and dark moths he counted in various regions of the countryside. These criticisms have led some critics of evolution to charge that the peppered moth story is "faked," or is "known to be wrong." Neither is true. In fact, the basic elements of the peppered moth story are quite correct. The population of dark moths rose and fell in parallel to industrial pollution, and the percentage of dark moths in the population was clearly highest in regions of the countryside that were most polluted. As Majerus, the principal scientific critic of Kettlewell's work wrote, "My view of the rise and fall of the melanic form of the peppered moth is that differential bird predation in more or less polluted regions, together with migration, are primarilty responsible, almost to the exclusion of other factors." (p. 155). So, what's going on here? Well, the best way to put it is that what we are seeing is the scientific process at its best. Majerus and other ecologists have carefully examined the details of Kettlewell's work and found them to be lacking. As Majerus explains, to be absolutely certain of exactly how natural selection produced the rise and fall of the carbonaria form, we need better experiments to show that birds (in a natural environment) really do respond to camoflage in the ways we have presumed, that the primary reason the dark moths did better in polluted areas was because of camoflage (and not other factors like behavior), and that migration rates of moths from the surrounding countryside are not so great that they overwhelm the influence of selection in local regions by birds. Until these studies are done, the peppered moth story will be incomplete. Not wrong, but incomplete. What we do know is that the rise and fall of dark-colored moths, a phenomenon known as "industrial melanism," remains a striking and persuasive example of natural selection in action. What we have to be cautious about is attributing 100% of the work of natural selection in this case to the camoflage of the moths and their direct visibility to birds." Ken Miller |
|
|
|
Mottled beauty
|
|
Close canopy |
Open canopy |
||||
|
|
melanic |
banded |
typical |
melanic |
banded |
typical |
|
Dyfed, 1984 |
3.4 |
19.0 |
77.6 |
0 |
0 |
100 |
|
Surrey, 1989 |
37.1 |
7.1 |
55.8 |
17.0 |
3.0 |
80.0 |
|
Surrey, 1990 |
53.7 |
8.5 |
37.8 |
17.8 |
6.7 |
75.5 |
|
Surrey, t991 |
39.8 |
9.2 |
51.0 |
21.0 |
5.7 |
73.3 |
|
Surrey, 1992 |
35.3 |
7.6 |
57.2 |
21.4 |
6.1 |
72.5 |
|
Surrey, 1993 |
39.2 |
8.5 |
52.3 |
19.5 |
6.3 |
74.3 |
|
Hants., 1991 |
21.1 |
7.9 |
71.0 |
6.4 |
2.4 |
91.2 |
|
Hants., 1992 |
19.7 |
11.1 |
69.2 |
9.2 |
3.0 |
87.8 |
|
Hants., 1993 |
18.2 |
10.1 |
71.7 |
8.2 |
3.2 |
88.7 |
|
Suffolk, 1991 |
28.4 |
7.2 |
64.4 |
9.7 |
5.2 |
85.2 |
|
Suffolk, 1992 |
32.5 |
6.4 |
61.1 |
16.4 |
4.9 |
78.7 |
|
Suffolk, 1993 |
26.7 |
6.1 |
67.2 |
13.5 |
4.2 |
82.3 |
|
Dyfed, 1991 |
1.6 |
16.3 |
82.2 |
0.9 |
9.1 |
90.0 |
|
Dyfed, 1992 |
4.7 |
21.4 |
73.9 |
1.2 |
7.2 |
91.6 |
|
Dyfed, 1993 |
1.9 |
18.9 |
79.2 |
0 |
7.4 |
92.6 |
|
Average % |
24.22 |
11.81 |
64.77 |
10.81 |
4.96 |
84.25 |
Grey pine carpet
|
|
Close canopy |
Open canopy |
||||
|
|
melanic |
banded |
typical |
melanic |
banded |
typical |
|
Glos., 1991 |
42.9 |
32.2 |
24.9 |
34.8 |
19.9 |
45.3 |
|
Suffolk, 1991 |
51.2 |
28.7 |
20.1 |
43.9 |
21.7 |
34.4 |
|
Suffolk, 1992 |
53.4 |
31.7 |
14.8 |
37.5 |
23.2 |
39.4 |
|
Suffolk, 1993 |
61.7 |
29.4 |
8.9 |
39.6 |
22.7 |
37.7 |
|
Average % |
52.3 |
30.5 |
17.18 |
38.95 |
21.88 |
39.2 |
Willow beauty
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Surrey, 1989 |
33.9 |
66.1 |
15.8 |
84.2 |
|
Surrey, 1990 |
35.5 |
64.5 |
6.0 |
94.0 |
|
Surrey, 1991 |
34.2 |
65.8 |
12.4 |
87.6 |
|
Surrey, 1992 |
43.7 |
56.3 |
14.8 |
85.2 |
|
Surrey, 1993 |
32.4 |
67.6 |
16.3 |
83.7 |
|
Hants., 1991 |
21.1 |
78.9 |
7.2 |
92.8 |
|
Hants., 1992 |
24.7 |
75.3 |
9.4 |
90.6 |
|
Hants., 1993 |
20.1 |
79.9 |
11.1 |
88.9 |
|
Suffolk, 1991 |
30.5 |
69.5 |
14.1 |
85.9 |
|
Suffolk, 1992 |
32.5 |
67.5 |
11.7 |
88.3 |
|
Suffolk, 1993 |
34.1 |
65.9 |
18.4 |
81.6 |
|
Dyfed, 1991 |
8.2 |
91.8 |
1.3 |
98.7 |
|
Dyfed, 1992 |
6.4 |
93.6 |
5.2 |
94.8 |
|
Dyfed, 1993 |
9.1 |
90.9 |
3.9 |
96.1 |
|
Average % |
26.17 |
73.83 |
10.54 |
89.46 |
Tawny-barred angle
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Dyfed, 1984 |
76.9 |
23.1 |
12.5 |
87.5 |
|
Hants., 1991 |
8.4 |
91.6 |
0 |
100 |
|
Hants., 1992 |
7.2 |
92.8 |
0 |
100 |
|
Dyfed, 1991 |
64.8 |
35.2 |
18.2 |
81.8 |
|
Dyfed, 1992 |
70.2 |
29.8 |
14.1 |
85.9 |
|
Dyfed, 1993 |
61.9 |
38.1 |
9.3 |
90.7 |
|
Average % |
48.23 |
51.77 |
9.02 |
90.98 |
Pale brindled beauty
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Cambs., 1991 |
28.4 |
71.6 |
16.3 |
83.8 |
|
Cambs., 1992 |
21.5 |
78.5 |
17.1 |
82.9 |
|
Cambs., 1993 |
30.2 |
69.8 |
19.2 |
80.7 |
|
Hants., 1991 |
18.8 |
81.2 |
13.5 |
86.5 |
|
Hants., 1992 |
14.1 |
85.9 |
13.9 |
86.0 |
|
Average % |
22.6 |
77.4 |
16 |
83.98 |
Dotted border
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Hants., 1989i |
12.7 |
87.3 |
1.5 |
98.5 |
|
Hants., 1989ii |
14.3 |
85.7 |
2.9 |
97.1 |
|
Hants., 1991 |
13.1 |
86.9 |
3.1 |
96.9 |
|
Hants., 1992 |
11.0 |
89.0 |
1.2 |
98.8 |
|
Cambs., 1991 |
9.2 |
90.9 |
2.0 |
98.0 |
|
Cambs., 1992 |
8.6 |
91.4 |
2.8 |
87.2 |
|
Cambs., 1993 |
10.4 |
89.6 |
1.4 |
98.6 |
|
Average % |
11.33 |
88.69 |
2.13 |
96.44 |
Common pug
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Cambs., Aug., 1991 |
86.9 |
13.1 |
79.3 |
20.7 |
|
Cambs., Spring, 1992 |
92.2 |
7.8 |
75.6 |
24.4 |
|
Cambs., Aug., 1992 |
88.7 |
11.3 |
80.6 |
19.4 |
|
Average % |
89.27 |
10.73 |
78.5 |
21.5 |
Engrailed
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Cambs., 1991 |
15.2 |
84.8 |
4.9 |
95.1 |
|
Cambs., 1992 |
19.8 |
80.2 |
7.3 |
92.7 |
|
Dyfed, 1992 |
26.8 |
73.2 |
14.2 |
85.8 |
|
Dyfed, 1993 |
31.2 |
68.8 |
12.7 |
87.3 |
|
Surrey, 1992 |
39.3 |
60.7 |
16.7 |
83.3 |
|
Average % |
26.46 |
73.54 |
11.16 |
88.84 |
Small engrailed
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Surrey, 1991 |
23.1 |
76.9 |
15.8 |
84.2 |
|
Surrey, 1992 |
29.4 |
70.6 |
12.7 |
87.3 |
|
Surrey, 1993 |
21.7 |
78.3 |
13.2 |
86.8 |
|
Cambs., 1991 |
18.6 |
81.4 |
7.3 |
92.7 |
|
Cambs., 1992 |
21.4 |
78.6 |
6.4 |
93.6 |
|
Dyfed, 1992 |
7.2 |
92.8 |
8.1 |
91.9 |
|
Dyfed, 1993 |
6.4 |
93.6 |
0 |
100 |
|
Average % |
18.26 |
81.74 |
9.07 |
90.93 |
Riband wave
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Surrey, 1990 |
34.6 |
65.4 |
11.5 |
88.5 |
|
Surrey, 1991 |
31.4 |
68.6 |
17.1 |
82.9 |
|
Surrey, 1992 |
27.7 |
72.3 |
15 |
85 |
|
Surrey, 1993 |
25.9 |
74.1 |
18.2 |
81.8 |
|
Cambs., 1992 |
26.7 |
73.3 |
21.4 |
78.6 |
|
Cambs., 1993 |
29.4 |
70.6 |
17.9 |
82.1 |
|
Average % |
29.28 |
70.72 |
16.85 |
83.15 |
Common marbled carpet
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Glos., 1991 |
44.6 |
55.4 |
22.9 |
78.1 |
|
Cambs., 1992 |
79.2 |
20.8 |
69.6 |
30.4 |
|
Cambs., 1993 |
75.8 |
24.2 |
69.2 |
30.8 |
|
Average % |
66.53 |
33.47 |
53.9 |
46.43 |
Pine carpet
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Glos., 1991 |
43.8 |
56.2 |
30.2 |
69.8 |
Large yellow underwing
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Glos., 1991 |
59.1 |
40.9 |
0 |
100 |
|
Cambs., 1992 |
46.2 |
53.8 |
41.7 |
58.3 |
|
Average % |
52.65 |
47.35 |
20.85 |
79.15 |
Lunar underwing
|
|
melanic (closed) |
non-melanic (closed) |
melanic (open) |
non-melanic (open) |
|
Cambs., 1991 |
15.5 |
84.5 |
8.4 |
91.6 |
|
Cambs., 1992 |
18.1 |
81.9 |
8.7 |
91.3 |
|
Average % |
16.8 |
83.2 |
8.55 |
91.45 |
|
Evolutionists arguments and rebuttals |
Evolutionist argument:
Evolutionists have little to say.